Hoofdstuk 4 Het holistische beeld
Laatste actualisatie: juli 2024 - Beperkte tekstuele aanvullingen
Eerste versie: september 2019
Het oorspronkelijke estuarium Grevelingen heeft zich in een serie beheersstappen ontwikkeld tot een zout meer, met beperkte uitwisseling van water met de Noordzee, en recentelijk ook met de Oosterschelde. Zie verder de Delta Expertise wiki
Figuur 4.1: Overzicht van de belangrijkste beheersstappen voor de uitwisseling van water uit het Grevelingenmeer met de Noordzee en de Oosterschelde van 1958 t/m 2023 (nog steeds actueel).
In dit hoofdstuk wordt het systeem beschreven vanaf ongeveer 1970, omdat na die tijd veel data beschikbaar zijn. Hiermee kan een beeld worden verkregen van het effect van het ’s zomers openen van de Brouwersdam (1980), en van de verschuiving van zomeropening naar het jaarrond open zijn (1999). In 2006 is de jaarrond opening volledig ingegaan. Het effect van de Flakkeese spuisluis, die recent weer is geopend, is waarschijnlijk klein en er wordt hier aangenomen dat dit verwaarloosd kan worden voor de algemene waterkwaliteit, alhoewel er wel positieve effecten werden gezien op vissen en daarmee het voedselweb van de Grevelingen.
De basis hiervoor is de effectketen zoals deze is opgesteld voor het Grevelingenmeer (figuur 4.2)

Figuur 4.2: Verrijkte effectketen voor het Grevelingenmeer. Bron: Delta expertise wiki.
4.1 Relaties hydrologie
Figuur 4.3: Overzicht van effectketen voor hydrologie
Waterbalans en uitwisseling Hier moet nog meer tekst komen over de wateruitwisseling
Stratificatie van het Grevelingenmeer treedt op in de zomermaanden (delta-expertise wiki 1 en Wijsman (2002)). De mate waarin dit optreedt hangt samen met de hydrologie. De aanvoer van zoet water uit polders en door neerslag veroorzaakt in rustige perioden een zoutstratificatie, waardoor in de zomer de oppervlaktelaag sneller opwarmt en er een sterke temperatuurstratificatie ontstaat (Wijsman 2002). Het jaarrond openstellen van de Brouwersdam in 1999 heeft er, door de grotere uitwisseling van water met de Noordzee, voor gezorgd dat de stratificatie minder sterk is geworden in de zomermaanden. Dit is in dit rapport uitgedrukt in de energie die nodig is per volume om de waterkolom geheel door te mengen (Boer, Pietrzak, and Winterwerp 2008) (figuur 6.25). De stratificatie is afgenomen doordat het bodemwater door deze menging een hogere temperatuur kreeg dan voor 1999. Hoewel de sterkte van de stratificatie na 1999 is afgenomen, is deze toch nog elk jaar in de zomer aanwezig. Ondanks de verminderde stratificatie is er dus nog steeds een barriëre voor het transport van zuurstof van de oppervlakte naar de diepere waterlagen. De verhoogde temperatuur in het bodemwater veroorzaakt een hogere afbraak van organisch materiaal, en daardoor een hogere zuurstofvraag. Hierdoor is de problematiek van zuurstofloos diep water niet afgenomen hoewel toch de stratificatie is verminderd.
4.2 Primaire productie
Figuur 4.4: Overzicht van effectketen voor primaire productie
Nutriëntenbeschikbaarheid
De waterkwaliteit in Grevelingenmeer kan aan de hand van een effectketen geanalyseerd worden. Mogelijke verbanden tussen hydrologie, biogeochemie in de diepere waterlaag, en primaire productieprocessen in de fotische zone zoals te zien in figuur 4.5 worden ingevuld door een beschrijvende analyse van de bestaande metingen.
Figuur 4.5: Effectketen voor beschikbaarheid van nutrienten voor fytoplankton
Lange termijn nutriëntenconcentraties
Voor de onderstaande analyse zijn jaargemiddelden bepaald door de gemeten waarden eerst per maand te middelen, en daarna per jaar, waarbij voor de winterwaarden alleen de maanden buiten het groeiseizoen (januari, februari, november, december) zijn beschouwd. De concentratie is dan een maat voor de totale hoeveelheid voor fytoplankton bruikbare nutriënten en geeft een potentieel aan voor de te verwachten voorjaarsbloei.
De resultaten voor de verschillende stations worden getoond. Er is helaas niet 1 station waarbij de hele tijdreeks beschikbaar is, en de variatie tussen de stations is, hoewel meestal erg klein, niet altijd te verwaarlozen.
Over de hele periode 1970 tot 2018 is vooral de concentratie van fosfaat sterk afgenomen. Deze afname loopt gelijk met een toename van het zoutgehalte (en dus verdunning met zeewater) vanaf 1978 (opening Brouwersdam), en wordt verder (en waarschijnlijk vooral) verklaard door maatregelen die de fosfaatbelasting vanaf land in het Rijn-Maasstroomgebied beperkten. In dezelfde periode is ook de Silicaatconcentratie afgenomen. Net als fosfaat, komt een groot deel van het silicaat van land of zoetwaterbronnen en de afname kan dus verklaard worden door een grotere uitwisseling met de Noordzee vanaf 1978.
De laatste jaren (vanaf 2011) neemt de zomerconcentratie voor fosfaat, ammonium en silicaat weer licht toe. Het is niet duidelijk waardoor dit komt. Een mogelijke oorzaak is een verhoogde nalevering vanuit het sediment, misschien ook gedreven door de hogere temperatuur van het bodemwater in de zomer, maar dit moet nog beter onderzocht worden.
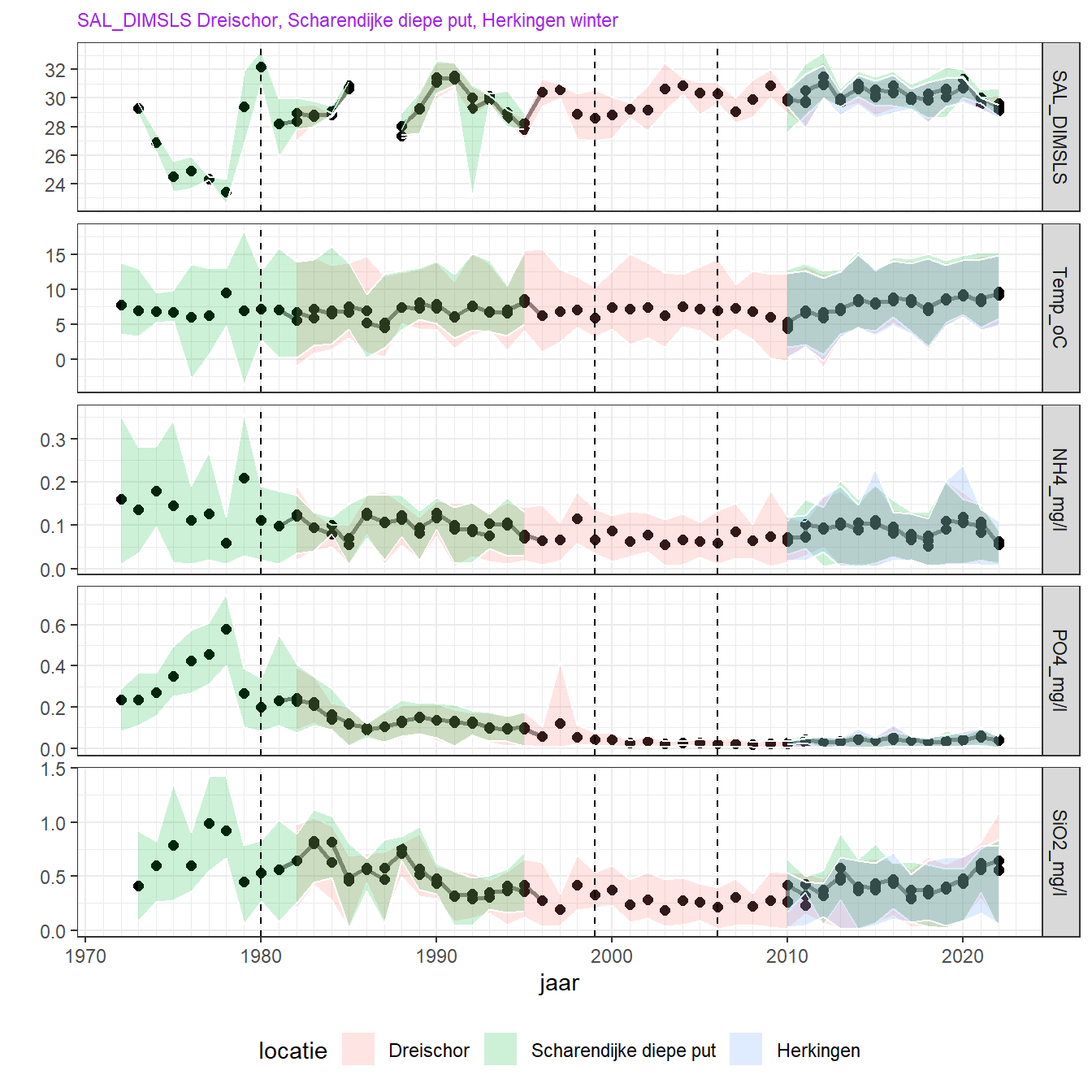
Figuur 4.6: Wintergemiddelde voor beschikbare metingen voor nutriëntenconcentraties aan de oppervlakte. De verticale lijnen geven de jaren 1980, 1999 en 2006 aan, toen veranderingen in het peilbeheer plaatsvonden.
Seizoensvariatie van nutriënten (sectie 8.2)
Een gemiddeld seizoen (vanaf 2000) laat zien dat een relatief korte en lage (8 ug/l chlorofyl-a) voorjaarsbloei gevolgd wordt door een afname in maart, waarschijnlijk doordat fosfaat beperkend wordt voor de groei. Silicaat is dan ook al erg laag. Een korte periode met laag chlorofyl wordt dan vanaf juni gevolgd door een lange periode (mei/juni - september) waarbij fosfaat niet meer beperkend is, en chlorofyl ongeveer even hoog is als gedurende de voorjaarsbloei. Gedurende de groeiperiode in de zomer is het anorganisch stikstof erg laag (waarschijnlijk beperkend) en is er silicaat in overvloed. Fosfaat en silicaat bereiken de hoogste concentraties gedurende de late zomer/herfst. Dit is te verklaren door remineralisatie van fosfaat en silicaat tijdens de warme periode in de zomer, waarbij het organisch materiaal van de voorjaarsbloei gedeeltelijk weer terugkomt in het water. Ook kan er in die periode teruglevering plaatsvinden van al eerder in het sediment opgeslagen nutriënten. Met name voor fosfaat is dit belangrijk, omdat fosfaat aan geoxideerd ijzer in de bodem wordt vastgelegd. Bij zuurstofloosheid in het sediment wordt ijzer gereduceerd en komt het fosfaat vrij. Zuurstofloosheid in het sediment komt ook voor als er nog zuurstof in de waterkolom aanwezig is. In het zomerhalfjaar wordt de zuurstofrijke bovenlaag van de bodem, die over het algemeen enkele centimeters dik kan zijn en sterk door bioturbatie wordt beïnvloed, dunner door hogere temperatuur waardoor de mineralisatiesnelheid toeneemt en door het gesedimenteerde, makkelijk afbreekbare verse organisch materiaal. De zuurstofrijke bovenlaag kan afnemen tot enkele millimeters of zelf helemaal verdwijnen. Hoeveel fosfaat dat gebonden is aan zwevend stof, vrij kan komen als het zwevend stof in zuurstofloze condities terecht komt, is niet duidelijk.
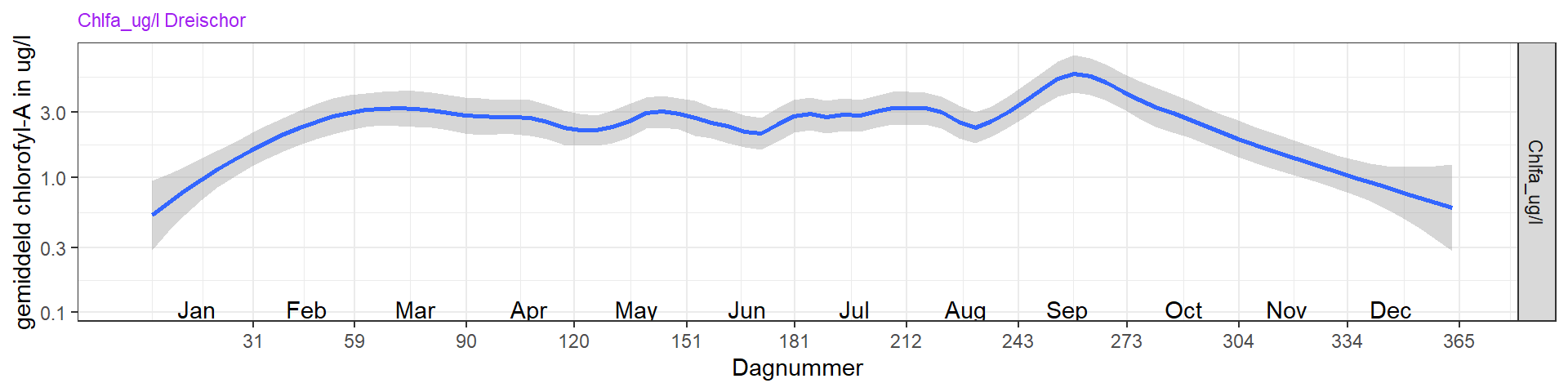
Figuur 4.7: Variatie van chlorofyl-a over het seizoen voor de jaren 2000 t/m 2022 op station Dreischor.
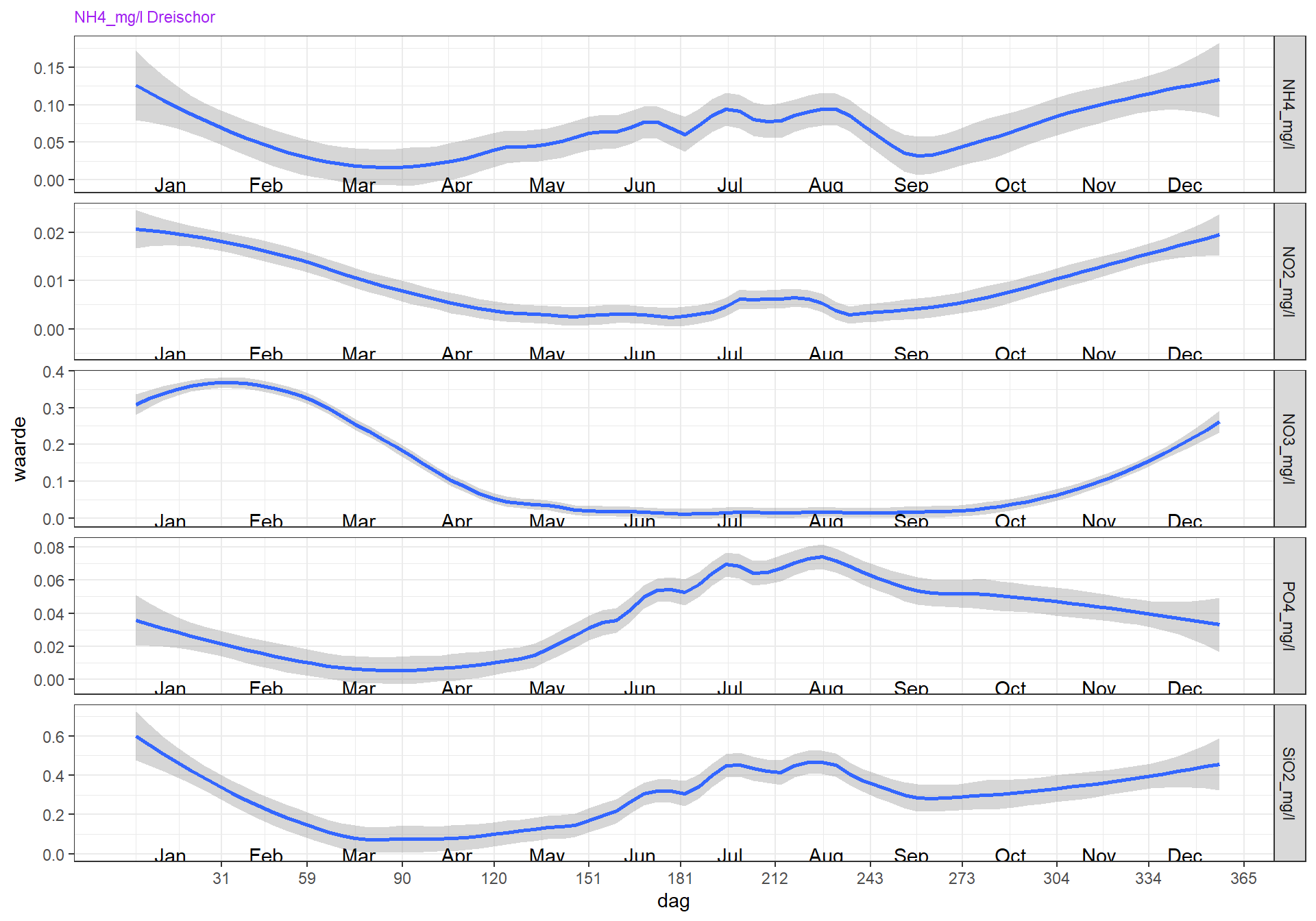
Figuur 4.8: Variatie van nutriëntenconcentratie over het seizoen voor de jaren 2000 t/m 2022 en station Dreischor.
Nutriëntratio’s
Ratio’s van nutriënten (sectie 8.2) vertellen iets over welk nutriënt limiterend kan worden wanneer er genoeg licht beschikbaar is. De optimale ratio voor fytoplankton die vaak aangehouden wordt is de zogenaamde Redfield ratio. Deze bedraagt C:Si:N:P = 106:15:16:1 mol/mol of C:Si:N:P = 41 : 14 : 7 : 1 gram/gram. Een grote verandering in de verhouding waarin nutriënten beschikbaar zijn kan leiden tot de groei van andere soorten fytoplankton. Als er bijvoorbeeld een tekort is aan silicaat, zullen er minder diatomeeën groeien, en meer flagellaten. Dit wordt over het algemeen als negatief beschouwd, omdat diatomeeën beter voedsel zijn voor zoöplankton en macrobenthos.
De wintergemiddelde ratio DIN/DIP is sinds de jaren 1970 toegenomen van zeer laag, naar ongeveer 10 met soms uitschieters tot 30 gN/gP. Dat is ruim hoger dan de Redfield ratio, wat impliceert dat de primaire productie in het voorjaar waarschijnlijk niet door stikstof wordt beperkt, wat overeen komt met het verloop van fosfaatconcentratie. Deze werd veroorzaakt door de sterke afname van winter fosfaatconcentraties in verhouding tot de bescheiden afname van stikstofcomponenten. De effecten van veranderingen in spuibeleid vallen hierbij in het niet. De laatste jaren lijkt weer een afname van winter DIN/DIP op te treden. Deze verandering begint enkele jaren nadat de Brouwersdam het gehele jaar open is gezet. Het zou kunnen dat hier een causaal verband is, namelijk een grotere invloed van het Noordzeewater op de concentraties in de Grevelingen (moet nog checken wat winter DINDIP in het kustwater is!).
De wintergemiddelde ratio van stikstof en silicaat (winter DIN/Si) vertoont een soortgelijke trend en is toegenomen van 0.5 naar 1
De wintergemiddelde ratio van fosfaat en silicaat (winter PO4/Si) is sterk afgenomen van 0.5 tot ongeveer 0.1 tegenwoordig. Dit is veel hoger dan de Redfield ratio (1/14), zodat de voorjaarsbloei wordt beëindigd met een overschot aan fosfaat.
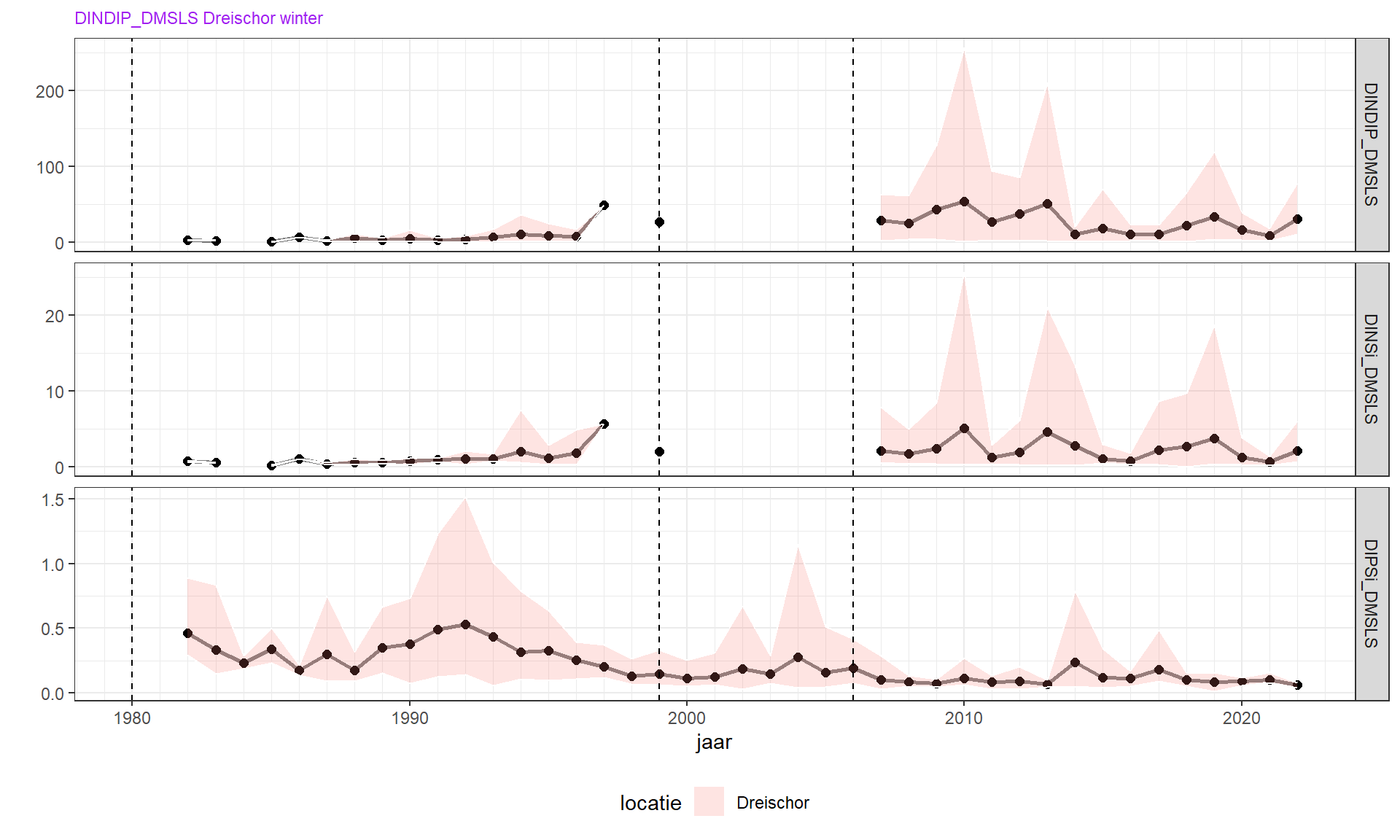
Figuur 4.9: Wintergemiddelde nutriëntenratio’s DIN/DIP, DIN/Si, en DIP/Si.
In de zomer lijkt er, gebaseerd op de concentraties, een overschot te zijn aan fosfaat en silicaat. Dit is terug te zien in lage DIN/DIP en DIN/Si ratio’s. in de zomer is ook de DIP/Si ratio relatief hoog. Door het overschot aan fosfaat en silicaat treedt echter normaal geen Si limitatie op.
Van ruwweg 2010 t/m 2013 zijn zeer hoge DIN/DIP en DIN/Si ratio’s gemeten. Dit valt samen met een relatief klein oppervlak waaronder het water zuurstofarm is.
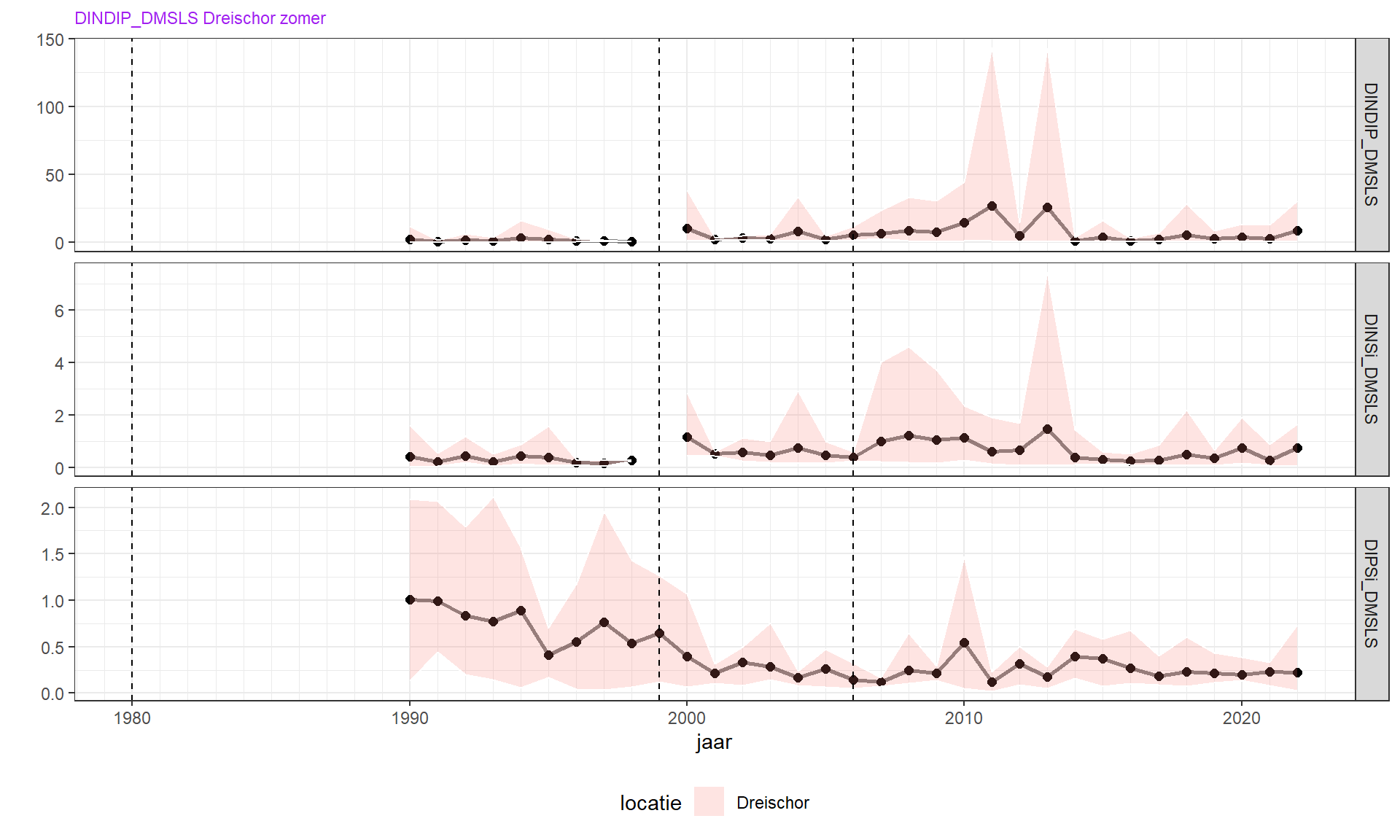
Figuur 4.10: Zomergemiddelde van nutriëntenratio’s. De periode van voor 1990 is opzettelijk weggelaten om een betere schaalverdeling te krijgen van DIP/Si
4.3 Lichtuitdoving in de waterkolom.
De hoeveelheid beschikbaar licht bepaalt, samen met de nutriëntenbeschikbaarheid, de primaire productie in de Grevelingen. Uitdoving van licht kan zorgen voor een lagere productie, en een slechtere benutting van de beschikbare nutriënten door fytoplankton. Aan de andere kant draagt het fytoplankton ook bij aan de lichtuitdoving. Lichtuitdoving wordt voornamelijk veroorzaakt door zwevend stof (meestal bestaand uit slib), opgelost organisch materiaal, en dus fytoplankton zelf. Wanneer licht beperkend is voor de groei van fytoplankton, wordt een negatief verband gevonden tussen de lichtuitdoving en fytoplankton of chlorofyl-a. Wanneer nutriënten limiterend zijn, en er voldoende fytoplanktonbiomassa is, wordt een positief verband gevonden.
Figuur 4.11: Effectketen voor doorzicht
De lichtuitdoving kan direct bepaald worden als extinctiecoëfficiënt, of in de vorm van transparantie gemeten worden als Secchidiepte (“Sechi_dm”). Deze geven allebei een indruk van hoe diep licht in de waterkolom doordringt, maar zijn omgekeerd evenredig. Voor de Grevelingen zijn geen extinctiecoëfficiëntwaarden bekend van voor 2002, maar wel secchidieptes. Er is hier voor gekozen om alleen de Secchidieptes mee te nemen in de analyse.
Voor de onderstaande analyse zijn de gemeten waarden eerst gemiddeld per maand, en daarna gemiddeld per jaar, waarbij alleen de maanden in het groeiseizoen (maart - oktober) zijn geselecteerd
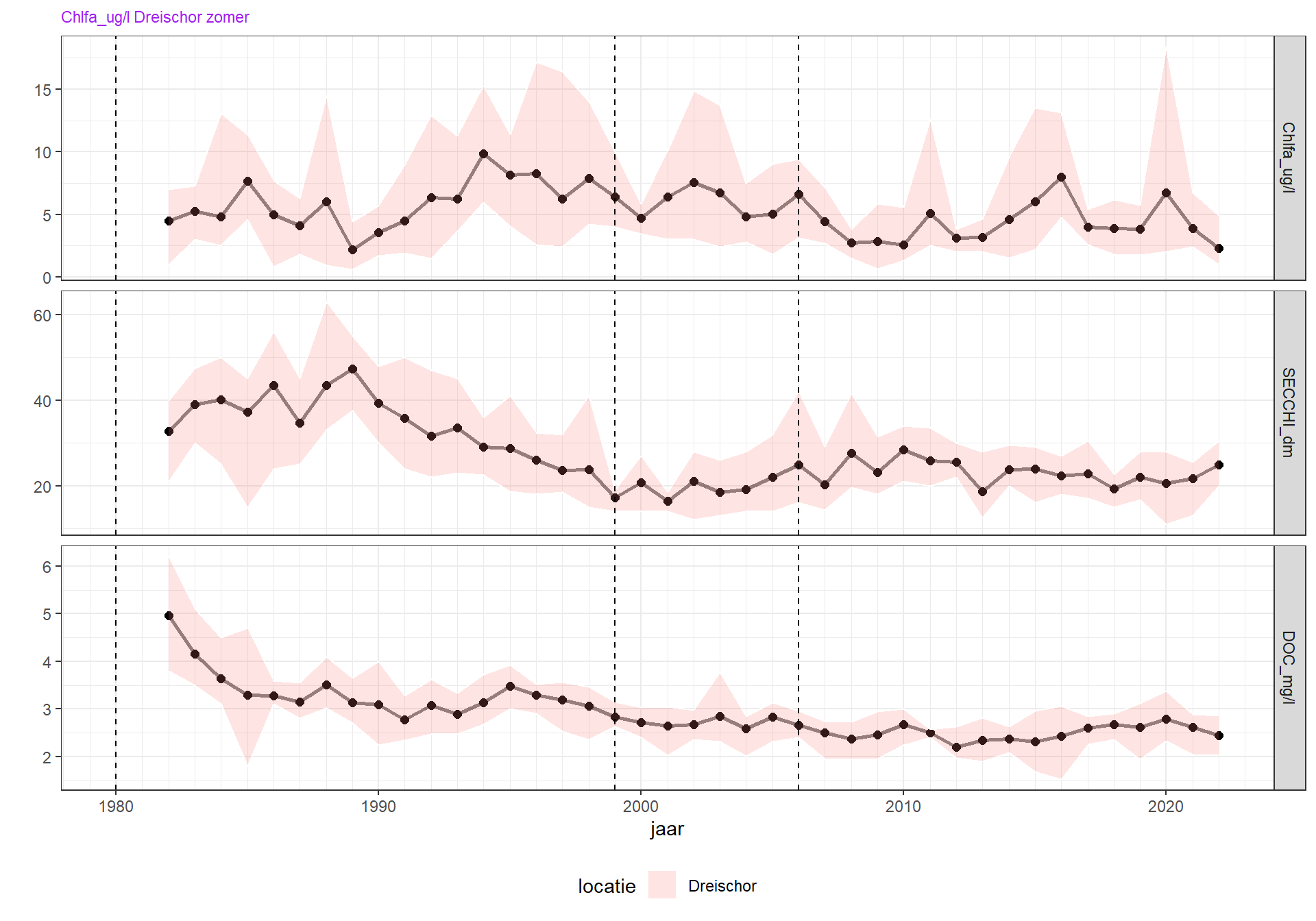
Figuur 4.12: Zomergemiddelden (maart-oktober) van doorzicht (als Secchidiepte) en verklarende variabelen DOC, zwevende stof en chorofyl-a over de hele periode waarin metingen beschikbaar zijn.
In de Grevelingen is een opvallende afname, grofweg een halvering, van de Secchidiepte tussen 1989 en 1999. Het Grevelingenmeer is dus troebeler geworden in die periode. Er lijkt over dezelfde tijd geen grote toename te zijn geweest van zwevend stof of DOC. Het is daarom niet waarschijnlijk dat de toename van de troebelheid veroorzaakt is door een verandering in zwevend stof of organisch materiaal. Wel is er een toename van het zomergemiddelde chlorofyl-a van ongeveer 5 ug/l in 1989 naar 10 ug/l in 1999 in die periode (figuur 4.12), en een toename van totaal koolstof in fytoplankton (figuur 4.13). Het is daarom waarschijnlijk dat de toename van de troebelheid veroorzaakt is door een toename van de gemiddelde fytoplankton biomassa in de zomer. Dit suggereert dat primaire productie in die periode overwegend nutriëntgelimiteerd was.
Het is niet duidelijk waarom fytoplankton toenam tussen 1989 en 1990. De veranderingen van chlorofyl-a en troebelheid die rond 1989 is ingezet, is lang nadat de Brouwersdam ’s winters werd opengezet (1980). Het is daarom onwaarschijnlijk dat dit een direct gevolg hiervan is. Het kan wel een indirect gevolg zijn, doordat meer nutriënten in het Grevelingenmeer hebben kunnen ophopen. In deze periode neemt de zomer DIP/Si ratio sterk af (figuur 8.14). Dit is consistent met een toename van diatomeeën tussen 1989 en 1999 (figuur 4.17) en zou de toename van de fytoplanktonbiomass kunnen verklaren.
Vanaf 1999 tot ongeveer 2010 neemt de Secchidiepte weer langzaam toe, en chlorofyl-a af. Het systeem wordt weer wat helderder. In 2013 is de tranparantie plotseling afgenomen. De laatste jaren neemt de transparatie weer toe. Deze variatie van de laatste jaren is niet goed te verklaren.
De uitdoving van licht door fytoplankton kan, behalve alleen met metingen van chlorofyl-a, ook onderzocht worden aan de hand van fytoplanktonbiomassa berekend uit tellingen. De totale fytoplankton biomassa (figuur 4.13) laat een gestage toename zien. Ook zijn er veranderingen in de verhouding tussen de fytoplankton groepen (figuur 4.17). Dit stemt deels overeen met de chlorofyl metingen, maar daarbij moet opgemerkt worden dat de biomassa /chlorofyl-a verhouding over de tijd sterk veranderd is (zie figuur 4.14). De toename in biomassa/chlorofyl-a duidt op een verschuiving van een meer lichtgelimiteerd systeem naar een meer nutriëntgelimiteerd systeem.
In een lichtgelimiteerd systeem kan verwacht worden dat fytoplantkonbiomassa correleert met het doorzicht (negatieve correlatie met totale extinctiecoefficient), maar in een nutriëntgelimiteerd systeem wordt dit niet verwacht. Als fytoplankton genoeg bijdraagt aan het doorzicht wordt hier zelfst een negatieve correlatie verwacht tussen biomassa en doorzicht.
De verandering in soortsamenstelling van fytoplanton en toename van biomassa ten opzichte van het chlorofyl-a gehalte zijn mogelijk de oorzaak van de afname van het doorzicht na 1990.
Deze metingen moeten echter met voorzichtigheid geanalyseerd worden aangezien de metingen van fytoplanton volume en biomassa van voor 2000 af kunnen wijken van de periode daarna door een wisseling van meetinstantie.
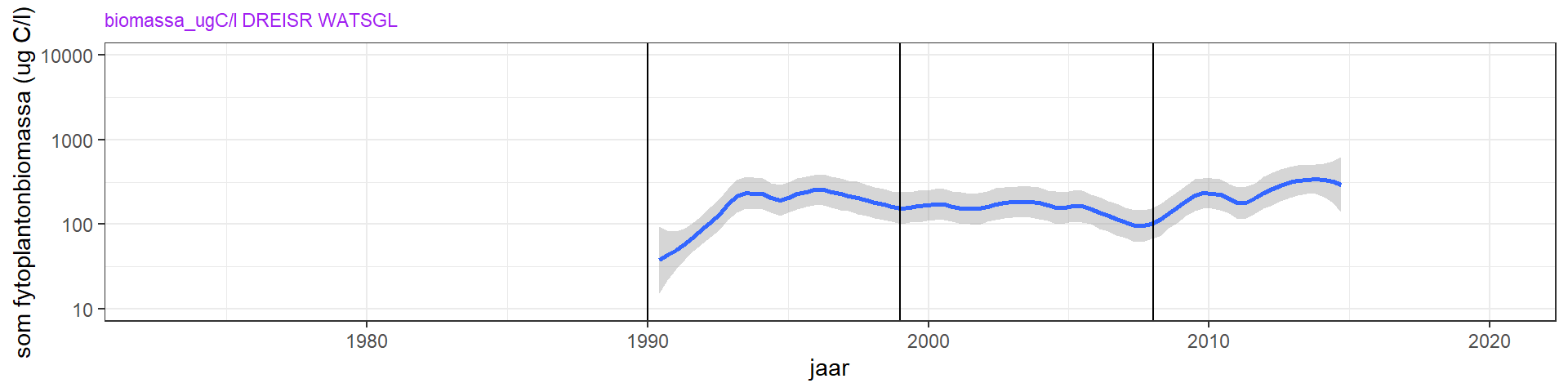
Figuur 4.13: Fytoplanktonbiomassa totaal
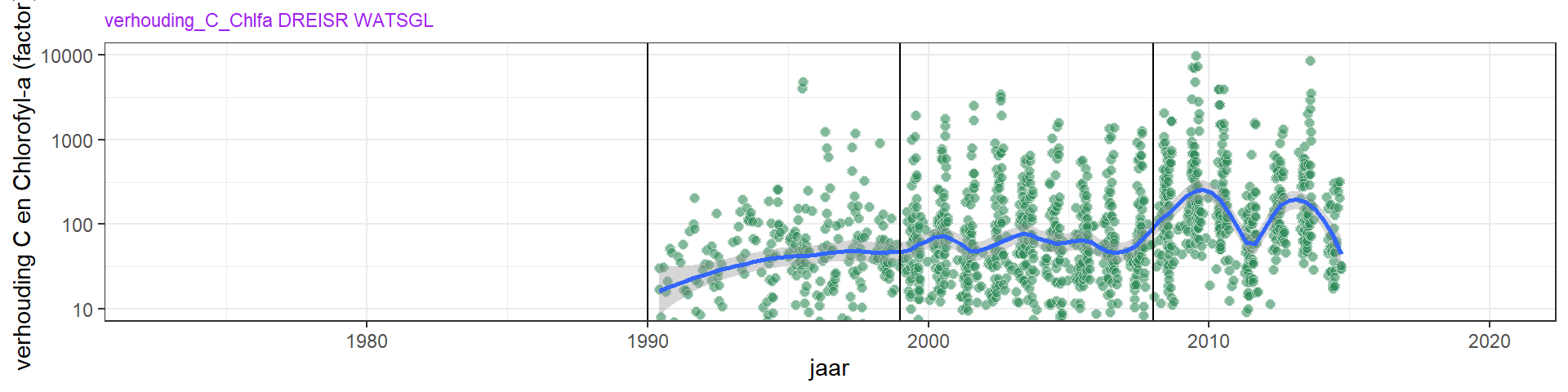
Figuur 4.14: De verhouding koostof/chlorofyl-a van fytoplankton in de Grevelingen.
4.4 Fytoplankton
De uitdoving van licht door fytoplankton kan, behalve alleen met metingen van chlorofyl-a, ook onderzocht worden aan de hand van fytoplanktonbiomassa berekend uit tellingen. De totale fytoplankton biomassa laat een toename zijn tussen 1990 en 1998. Dit komt overeeen met de toename in chlorofyl-a. Ook de hogere concentratie van chlorofyl-a in de laatste jaren (figuur 4.12) komt terug in de biomassametingen van fytoplankton (figuur 4.15).
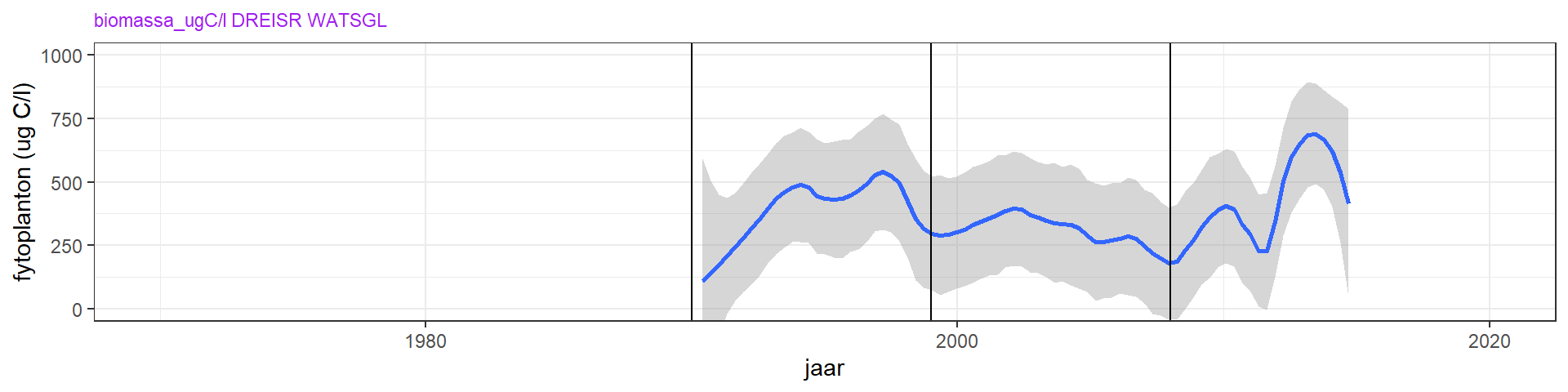
Figuur 4.15: Zomergemiddelde van de biomassa aan fytoplankton vanaf 1990. Er zijn geen tellingen van voor 1990. De blauwe lijn is een Loess local regression model, en de grijze band de standaardfout hiervan.
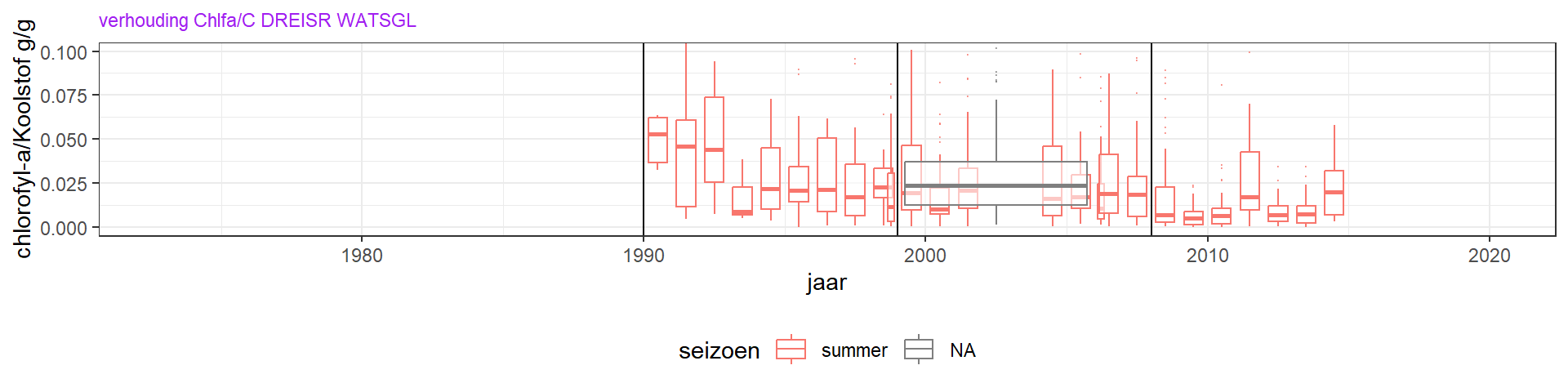
Figuur 4.16: De verhouding fytoplankton koolstof/chlorofyl-a in de Grevelingen.
De verhouding tussen gemeten chlorofyl-a en totale fytoplanktonbiomassa (chlfa/C) in de zomer (maart - oktober) geeft het chlorofylgehalt aan van het aanwezige fytoplankton (figuur 4.16). Hier zit erg veel variatie in per monster en jaar, door de variatie in chlorofyl-a concentratie maar vooral door de variatie in de fytoplanktontellingen en conversie naar koolstof. De chlfa/C verhouding varieert van rond de 0.4 begin jaren 1990, tot 0.2 of zelfs 0.1 in latere jaren. Waarden rond de 0.4 geven aan dat fytoplankton toen gelimiteerd werd door licht (Cloern et al. (1995), Wang et al. (2009)), en de lagere waarden duiden op nutriëntlimitatie. Ter indicatie, in de Noordzee varieert chlfa/C van ongeveer 0.1 in de kustwater tot 0.05 ver uit de kust (Alvarez-Fernandez and Riegman (2014)). Het lijkt er dus op dat de primaire productie vanaf 1993 nutriënt gelimiteerd is. Daarvoor (1990 - 1993) suggereert de chlfa/C verhouding licht limitatie. Voor 1990 zijn er geen fytoplanktontellingen uitgevoerd, waardoor hier geen ratio’s te berekenen zijn.
Ook zijn er veranderingen in de verhouding tussen de fytoplankton groepen. Tussen 1990 en 2000 waren andere soorten dan diatomeeën de dominante groep. Na 2000 is het aandeel van diatomeeën weer iets hoger, en vanaf 2015 zijn diatomeeën de dominante groep. Dit is consistent met de sterke daling van afname in DIN/Si vanaf 2015 (figuur 8.21).
De metingen van fytoplanktonbiomassa en soortensamenstelling moeten met enige voorzichtigheid genomen worden aangezien de metingen van fytoplanton volume en biomassa van vóór 2000 af kunnen wijken van de periode daarna door een wisseling van meetinstantie.
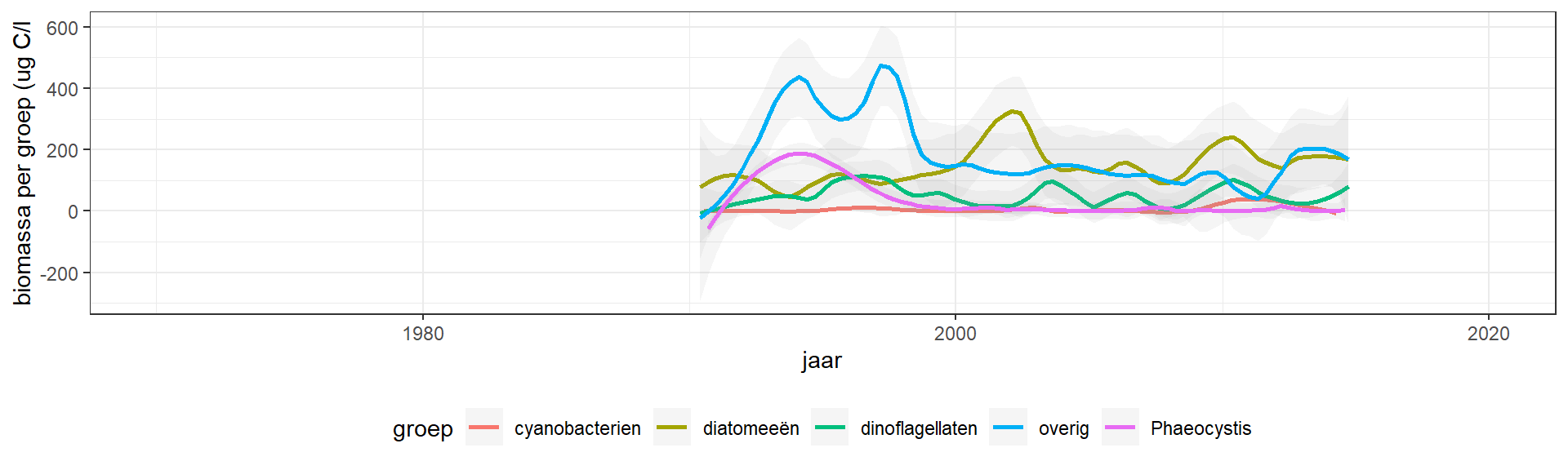
Figuur 4.17: Verloop van de groep fytoplanktonbiomassa in het Grevelingenmeer zoals gemeten op station Dreischor ug C/l aan de oppervlakte. Er zijn 4 metingen verwijderd omdat ze buiten de schaal vallen. Deze metingen zijn wel meegenomen in de curve fit.
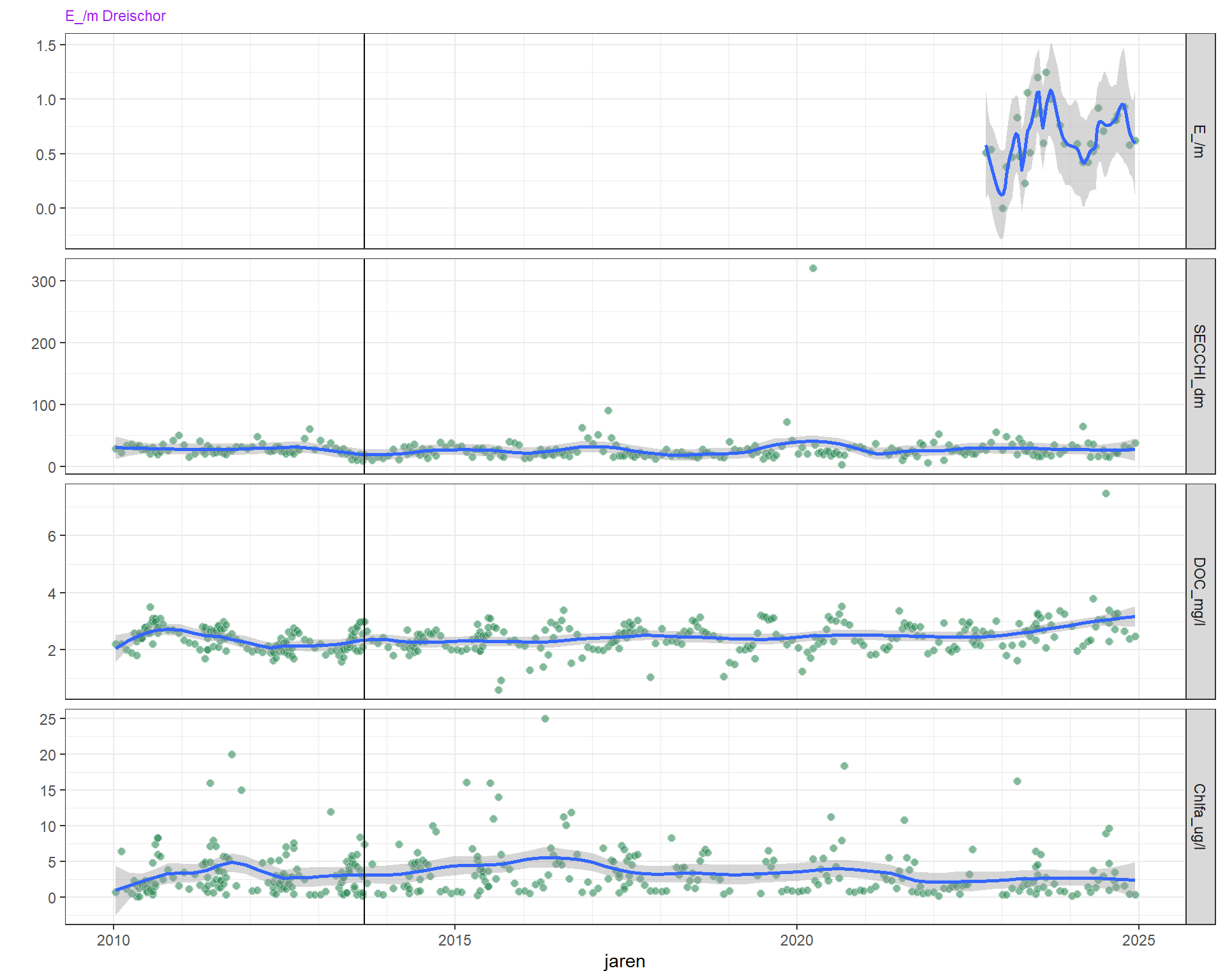
Figuur 4.18: Extinctiecoëfficiënt, doorzicht (secchi) en verklarende variabelen DOC, zwevende stof en chorofyl-a variatie over de laatste jaren. De verticale lijn geeft een jaar (2013) aan met erg weinig doorzicht
In recentere jaren is er variatie in de secchischijf diepte waarbij de diepte in de winter grosso modo iets hoger is dan in de zomer. Het is niet direct duidelijk waardoor dit veroorzaakt wordt, maar in de meeste jaren is het zwevend stof, chlorofyl en DOC verhoogd, wat samenvalt met een kleinere doorzichtdiepte. In de zomer van 2013 viel een zeer laag doorzicht niet samen met verhoogd zwevend stof, maar wel met verhoogd chlorofyl en opgelost koolstof.
4.5 Zuurstof in de diepere waterlagen
In de diepe putten van het Grevelingenmeer treedt in de zomer vaak zuurstofloosheid op (Wetsteijn 2011). Deze wordt veroorzaakt door de bijna permanente zoutstratificatie, dus beperkte uitwisseling van water in de verticaal. In de zomer is door de hogere temperatuur en beschikbare organische stof uit plankton de zuurstofvraag hoger dan de toevoer door verticale menging. Dit veroorzaakt in de verschillende putten zuurstofarm of zuurstofloos bodemwater in de zomer.
De zuurstofloosheid in de onderste waterlaag is hier bepaald door het oppervlakte in hectare te bepalen waarbij de diepere waterlaag een zuurstofconcentratie had die lager is dan 3 mg/l. Deze indicatorwaarde is bepaald van 1979 (het begin van de gtso metingen) tot en met 2018. Het totale oppervlak zuurstofarm water vertoont een toename over deze periode, maar ook veel variatie. De periode 2010 - 2013 waarbij zomer DIN/DIP hoog was, lijkt gedeeltelijk samen te vallen met een periode met een klein oppervlak zuurstofarm diep water. Het ontbreken van winterwaarden voor zuurstofarm oppervlakte ligt gedeeltelijk aan het meetprogramma. In de Zomer zijn veel meer metingen gedaan dan in de winter. Overigens treedt ’s winters veel minder vaak zuurstofarm water op.
Het oppervlakte zuurstofarm water verschilt per put, mede door verschillen in temperatuurstratificatie. Daar wordt hier nu niet dieper op ingegaan. Voor een uitgebreidere analyse, zie Wetsteijn (2011) en hoofstuk 8.4.
Een breekpuntanalyse van het oppervlakte zuurstofarm water (figuur 8.67) laat zien dat er waarschijnlijk in 1999 een verandering is opgetreden in het gemiddeld oppervlak met een zuurstofconcentratie lager dan 3 mg/l in de zomer. Dit valt samen met het veranderde spuibeleid, waardoor er een grotere verversing optrad, en er een grotere hoeveelheid zout water in de Grevelingen werd gelaten.
De toegenomen uitwisseling met de Noordzee heeft er toe geleid dat de sterkte van de stratificatie in de diepe putten is afgenomen (figuur 6.25 en sectie 6.4). Toch is hierdoor de problematiek met zuurstofarm bodemwater niet verminderd (figuur 8.65 en 8.66).
Verklarende factoren voor het feit dat het maximale zuurstofarm oppervlak toegenomen is bij de toegenomen jaarrond verversing door Noordzeewater zouden kunnen zijn:
- verhoogde invoer van organisch materiaal door verhoogde uitwisseling met de Noordzee
- verhoogde lokale productie en sedimentatie door verhoogde stikstofgehaltes die weer veroorzaakt zijn door een verhoogde uitwisseling met de Noordzee.
- verhoogde temperatuur van het bodemwater in de diepe putten, waardoor de afbraak van organisch materiaal is versneld, en er een snellere en grotere zuurstofvraag is ontstaan. Een voorwaarde hiervoor is dat er genoeg organisch materiaal voorhanden is.
Van deze factoren is alleen de laatste gemakkelijk met de meetdata te controleren. De regelmatige GTSO metingen in het Grevelingenmeer geven een duidelijke verandering van de bodemwatertemperatuur aan die gelijktijdig optreedt met de verandering van het spuibeleid (figuur 8.115). De toegenomen mengeing zorgt ervoor dat de verticale temperatuurgradiënt afvlakt, waardoor de temperatuur in de diepe lagen in de zomer sneller toeneemt en hoger wordt. Dit was ook al geconstateerd door Wetsteijn (2011) , maar kan nu ondersteund worden met meer meetjaren aan data, in combinatie met een breekpuntanalyse. Voor 1999 was de gemiddelde zomertemperatuur van het diepe water rond de 8 oC. Na 1999 is de gemiddelde zomertemperatuur van het diepe water rond de 12 oC. Deze temperatuurverandering treedt niet op in de oppervlaktelaag. Hoewel het aannemelijk is dat ook de saliniteit van het bodemwater is veranderd na 1999, kon dit niet met de data ondersteund worden. Het lijkt er meer op de dat de jaar tot jaar variatie minder groot is geworden door de vergrote uitwisseling (figuur 8.114).
4.6 Ecologie Water
4.6.1 Macrobenthos
Uit de analyses blijkt dat de afsluiting van het Grevelingenmeer en het openen van de Brouwerssluis een sterke invloed hebben gehad op de samenstelling van het macrobenthos. Zo nam de soortenrijkdom na de sluiting met een kwart af en vonden forse verschuivingen plaats onder soorten die zich hadden weten te handhaven. Door het wegvallen van het getij konden bepaalde gebieden (20% van de open Grevelingen) gekoloniseerd worden. Deze gebieden werden onderhevig aan sterke stroming en sedimentbewegingen waardoor alleen specialisten zich daar konden vestigen. Na het openen van de Brouwerssluis in 1978 vonden verschillende systematische veranderingen plaats: * Het inlaten van zouter Noordzeewater leidde in het eerste jaar tot stratificatie waardoor grote sterfte van het macrobenthos plaatsvond beneden de 8 m; * Door ophoping van organisch materiaal in de diepere delen van het meer door het wegvallen van het getij steeg de zuurstofbehoefte en verslechterde hierdoor de zuurstofomstandigheden nabij de bodem; * Schelpen en andere harde ondergronden raakten ondergesneeuwd door organisch materiaal waardoor broedval steeds minder succesvol werd. De verbinding met de Noordzee had ook positieve gevolgen omdat verdwenen soorten terugkeerden en de voedselsituatie zich verbeterde. Tussen 1990 en 2000 bleven zowel de totale gemiddelde biomassa als de gemiddelde aantallen gelijk maar bleef de soortensamenstelling veranderen. De biomassa in deze periode was ook vrij hoog en begon na 2000 sterk af te nemen alhoewel de biomassa vanaf 2010 weer iets toenam. Het Grevelingenmeer wordt gekenmerkt door een aantal exoten waarvan sommige een belangrijke rol spelen in het functioneren van het meer. Zo was het muiltje gedurende lange tijd de meest dominante soort in termen van biomassa, de laatste jaren is het belang van deze soort echter afgenomen en nu domineert de Japanse oester. Als biobouwers creëren deze soorten extra habitat voor heel wat andere soorten en als filterfeeders hebben ze een grote invloed op de benthisch-pelagische koppeling. Nu, vijftig jaar na de afsluiting van het Grevelingenmeer, is de macrobenthosgemeenschap nog steeds in ontwikkeling.
4.6.2 Vissen
Het algemene beeld is dat de afsluiting van het Grevelingenmeer en het sluisbeheer van de Brouwerssluis een sterke invloed hadden op de samenstelling van de vispopulatie. Als estuarium fungeerde de Grevelingen vooral als paai-, opgroei- en foerageer habitat voor mariene en estuariene soorten of als doortrek gebied naar de rivieren voor migrerende soorten. De aanleg van de Grevelingendam en Brouwersdam hadden tot gevolg dat vissen werden opgesloten in het ontstane Grevelingenmeer en voor sommige soorten voortplanting belemmerd werd omdat er geen doortrek naar de rivieren mogelijk was, soorten hun paaigronden in zee niet konden bereiken en er geen aanwas van larven plaatsvond. Na de afsluiting is het vermogen tot voorplanting bepalend geweest voor de ontwikkeling van de visfauna in het Grevelingenmeer, met als gevolg een daling in de diversiteit en een verandering in de samenstelling van de visfauna van voornamelijk grote naar kleine soorten. De opening van de Brouwerssluis in 1978 maakte uittrek naar paaigronden, uitwisseling van larven en intrek van zomergasten mogelijk. Echter belemmerde de zomersluiting (april-september) vanaf 1980 intrek van zomergasten en deels ook uitwisseling van larven in het voorjaar afhankelijk van het moment van sluiting en moment van de larven piek. Het relatief intensieve bemonsteringprogramma tussen 1960 en 1994 heeft duidelijke relaties kunnen leggen tussen het spuibeheer en de samenstelling van de vispopulatie. Echter, rondom de aanpassing van het sluisbeheer in 1999 en 2005 hebben geen bemonsteringen plaatsgevonden met een boomkor waardoor de ontwikkelingen moeilijker te bepalen waren. Aan de hand van duikwaarnemingen is aangegeven dat het jaarrond openstellen van de Brouwerssluis positieve effecten heeft gehad op de diversiteit, o.a. doordat seizoensgebonden trekgedrag ook in de zomer weer mogelijk was. Daarnaast werden ook Noordzee vissoorten in grotere hoeveelheden gevangen en verjongde de scholpopulatie doordat uitwisseling van larven verbeterde. In de huidige situatie wordt de visstand gekenmerkt door redelijke aantallen maar een lage biomassa doordat het vooral kleine vissen betreft. Sinds de opening van de Flakkeese spuisluis (voorjaar 2017) wordt aangegeven dat het aantal soorten in de directe omgeving licht is toegenomen op basis van onderzoek in 2017.
4.6.3 Vogels
- Broedvogels. Van alle soorten broedvogels is er momenteel niet één die het voor het Grevelingenmeer gestelde Natura 2000 doelaantal haalt. Desondanks is het Grevelingenmeer nog steeds van groot belang voor kustbroedvogels in het Deltagebied. Het is landelijk het belangrijkste broedgebied voor de zeldzame strandplevier. Beheersmaatregelen zijn echter nodig om geschikt broedhabitat te behouden.
- Niet-broedvogels/Viseters. Verreweg de belangrijkste groep watervogels in het Grevelingenmeer zijn de viseters. Het Grevelingenmeer is / was voor middelste zaagbek, fuut, geoorde fuut en kuifduiker van groot internationaal belang. De trend van de meeste viseters is negatief en het jaargemiddelde van alle soorten, uitgezonderd de aalscholver, is lager dan het Natura2000 doel. De aantallen lijken voedsel-gerelateerd.
- Bodemdiereters/Steltlopers. Voor steltlopers is het Grevelingenmeer als foerageergebied van minder belang vanwege het geringe oppervlak foerageerhabitat. Vanwege de rust en ruimte zijn er enkele belangrijke hoogwatervluchtplaatsen voor steltlopers uit de Oosterschelde in het Grevelingenmeer.
- Bodemdiereters/Eenden. Voor bergeend en brilduiker is het Grevelingenmeer van relatief groot belang. De trend van de brilduiker is negatief maar dit wordt deels veroorzaakt door externe oorzaken.
- Planteneters. Belangrijke aantallen ganzen en eenden (planteneters) komen voor in het Grevelingenmeer. Ze foerageren op de buitendijkse graslanden en op de ondiepe oevers. De trend wijkt in de meeste gevallen niet af van de trend in het Deltagebied. Een uitzondering hierop zijn de meerkoet en knobbelzwaan. Beide soorten foerageren op onderwaterplanten en verdwenen bijna helemaal door afname van voedselplanten in het Grevelingenmeer.
- Roofvogels. Het Grevelingenmeer is broedgebied en overwinteringsgebied voor diverse soorten roofvogels. Alleen de bruine kiekendief (broedvogel) en slechtvalk (wintergast) zijn aangewezen als Natura 2000-soorten. Voor beide soorten geldt dat trend niet afwijkt van trend in het Deltagebied. Het aantal broedparen bruine kiekendief is lager dan het Natura 2000-doel. Het jaargemiddelde van de slechtvalk is stabiel en schommelt rond het Natura2000-doel.
- Zeezoogdieren. De gewone (vele tientallen) en de veel schaarsere grijze zeehond nemen in het Grevelingenmeer toe als uitvloeisel van de sterke toenames in de getijdewateren van het Deltagebied. Dit proces zal waarschijnlijk nog enige tijd doorzetten, al is sterfte als gevolg van verdrinking in fuiken mogelijk een beperkende factor.
Bron: WMR, 2019. Ecologisch onderzoek Getij Grevelingen (p122/123).
4.7 Ecologie - Land
- Na het droogvallen van het voormalige intergetijdengebied traden verschillende ontwikkelingen op (successie-reeksen), afhankelijk van (1) de uitgangssituatie (schor of zandplaat), (2) de hoogteligging, (3) de bodem (zandig of kleiïg) en (4) het gevoerde beheer.
- Naast successie zijn inklinking, erosie en het gevoerde peilbeheer van belang geweest voor de ontwikkeling van vegetatie.
- Om de ontwikkeling tot duinvallei-vegetaties goed te begrijpen zijn twee hydrologische systemen van belang: + (1) terreindelen waar slecht doorlatende klei- of veenlagen ondiep in de zandige ondergrond zitten (vaak op 1 tot 3 meter of zelfs minder). Deze locaties zijn dynamisch: erg nat in de winter, met stagnerend water, maar drogen sterk uit in de zomer, waarbij zelfs kans is op enige verzilting. Deze systemen zijn aanwezig op de Slikken Flakkee Noord en Zuid, en aan de oostzijde van de Hompelvoet. Hier zijn duinvallei-vegetaties minder goed ontwikkeld en hebben ze de neiging te verdrogen of te verzuren. Dit uit zich onder meer in een toename van kruipwilg, een hoog aandeel van soorten van wisselende waterstanden en lagere soortenrijkdom. Groenknolorchis komt hier bijvoorbeeld niet of nauwelijks voor. + (2) terreindelen met diepe, zandige ondergrond met een goed ontwikkelde zoetwaterbel in de ondergrond. Aan de randen van de zoetwaterbel treedt water uit op maaiveld en stroomt het af naar het meer. Dit type hydrologisch systeem is aanwezig op de Slikken Flakkee Midden, Markenje, het grootste deel van de Hompelvoet, de Veermansplaat (deze is iets slibrijker aan de zuidzijde), de Stampersplaat en Dwars in de Weg. Hier zijn duinvallei-vegetaties beter ontwikkeld en beter bestand tegen weersschommelingen (vernatting, verdroging) met stabiele aanvoer van kalkrijk grondwater. De verwachting is dat hier de duinvalleien bij het huidige beheer meerdere decennia kunnen standhouden.
- Op de oude schorren heeft zich binnen 10 jaar een ruigtevegetatie ontwikkeld op de kleibodem, een ontwikkeling die typerend is voor voedselrijke omstandigheden. Hoewel de soortensamenstelling van de ruigte wel verandert, vindt er niet of nauwelijks verdere successie plaats naar struweel of bos.
- Op de niet beheerde zandplaten is snel verzoeting opgetreden, waarbij successie plaats vond naar struweel en wilgenbos. Langs de oevers blijft onder zout invloed een smalle zone aanwezig van pioniervegetatie met zeekraal, zilt en brak grasland en duinvallei-vegetatie.
- Op de beheerde zandplaten (maaien/begrazen) wordt successie tegengegaan, en handhaaft zich op de meeste locaties een korte grasmat: duinvallei-begroeiingen met een hoog aandeel van zeggensoorten, die onder invloed staan van kalkrijk grondwater. Inmiddels zijn vrijwel alle beheerde delen verzoet, met uitzondering van de laagste oeverdelen waar nog steeds zilte en brakke (gezoneerd) delen worden aangetroffen, en de verzoeting nog enigszins doorgaat.
- De Groenknolorchis werd ontdekt rond 1990 en nam sindsdien gestaag toe met de grootste populatie op de Veermansplaat, en kleinere populaties op de Slikken Flakkee Zuid, Hompelvoet, Stampersplaat en Dwars in de Weg. Door seizoensinvloeden kunnen aantallen sterk fluctueren: na een aantal piekjaren zijn de aangetroffen aantallen na de droge jaren 2017 en 2018 sterk afgenomen. Desondanks herbergt het Grevelingenmeer nog steeds één van de grootste populaties van deze soort in ons land.
- De Noordse Woelmuis heeft zich na de afsluiting kunnen vestigen en komt wijdverspreid voor, echter met de hoogste dichtheden op de Slikken van Flakkee Noord en het vogeleiland Markenje.
Bron: WMR, 2019. Ecologisch onderzoek Getij Grevelingen (p126/127).