Hoofdstuk 9 Ecologie water
9.2 Fytoplankton
Fytoplankton is de basis voor het overige leven in het Grevelingenmeer. De ontwikkeling van de hoeveelheid algenbiomassa en de samenstelling van de fytoplanktongemeenschap over de seizoenen en de jaren bepaalt de draagkracht voor zoöplankton, benthos, vissen, vogels en zeezoogdieren. De variabiliteit van de algenbiomassa wordt vertegenwoordigd door de concentratie chlorofyl-A, en is besproken onder 8.8.
Hieronder wordt de samenstelling van het fytoplankton besproken aan de hand van de samenstelling van het fytoplankton als de concentratie van specifieke indicatorgroepen, omgerekend naar mg C per liter, zowel over seizoenen als over de jaren.
De grens tussen fototrofe algen zoals diatomeeën (die alleen van zonlicht en CO2 koolhydraten kunnen maken), en mixotrofe algen zoals dinoflagellaten (die ook in staat zijn in hun voeding te voorzien door bijvoorbeeld deeltjes op te nemen) is niet strikt, en kan op verschillende manieren worden ingevuld. De ontwikkeling van de verhouding tussen deze groepen kan vaak worden gerelateerd aan de trofiegraad (voedselrijkdom) van het Grevelingenmeer.
De aanwezigheid van plaagalgen is relevant voor o.a. de zwemwaterkwaliteit en de voedselveiligheid. Schelpdieren bedoeld voor consumptie kunnen voor de mens giftig worden als zij gifstoffen van plaagalgen opnemen in hun weefsel. Daarnaast is de concentratie van plaagalgen ook gerelateerd aan een onbalans in en/of verhoogde concentraties van voedingsstoffen in combinatie met oplopende watertemperaturen en mogelijk een indicator daarvoor. Als indicatorsoort voor plaagalgen is hier Phaeocystis (globosa) ngenomen. Deze soort wordt in Nederland als plaagalg gezien, niet vanwege gifstoffen (die produceert het niet), maar vooral vanwege de schuimvorming op het strand.
Chlorofyl-a en Fytoplanktonsamenstelling (incl. plaagalgen) De fytoplanktongemeenschap is voor deze analyse onderverdeeld in 4 hoofdgroepen: diatomeeën, dinoflagellaten, Phaeocystis en overig. Meetgegevens zijn beschikbaar van 1990 tot en met heden, meetfrequentie is ca. tweewekelijks, en er zijn op drie waterdieptes monsters genomen: aan het wateroppervlak, op de spronglaag (als aanwezig, en op variërende absolute diepte), en nabij de bodem. De monsters op de spronglaag en nabij de bodem zijn alleen genomen als er een spronglaag aanwezig was (anders is de aanname een gemengde waterkolom en weinig verloop van concentratie chlorofyl-a met de diepte); deze spronglaag, veroorzaakt door verschillen in saliniteit en temperatuur, is meestal alleen aanwezig in de zomermaanden.
In deze rapportage zijn fytoplanktongegevens vanaf 2000 verwerkt. Dit is een door RWS-CIV geharmoniseerde reeks. De gegevens van vóór 2000 kunnen niet op dezelfde manier geharmoniseerd worden.
De biomassa’s zijn vanuit tellingen omgerekend naar koolstof met behulp van sjabloonwaarden per soort. Niet voor alle soorten is zo’n sjabloonwaarde op dit moment beschikbaar. Hierdoor zullen de koolstofconcentraties per groep iets lager uitvallen. Er wordt door Bureau Waardenburg op dit moment gewerkt aan een completere lijst.
Voor chlorofyl-a figuren wordt verwezen naar sectie 8.8
Het verloop van de algenbiomassa in de oppervlaktemonsters over de jaren 1990 tot en met 2014 per groep (figuur 9.1) laat zien dat Phaeocystis in het begin van de 90er jaren sterk toenam, en een even groot aandeel in de algenbiomassa had als de diatomeeën en de dinoflagellaten. Vanaf 2000 is dit aandeel weer gedaald tot op 1 - 10% van de diatomeeënbiomassa, in absolute waarden tot rond de 1 tot 5 ug C/l. Diatomeeën en dinoflagellaten hebben een vergelijkbaar aandeel in de algenbiomassa, variërend tussen de 10 en 100 ug C/l, met wat lagere waarden voor 2000, en wat hogere daarna.
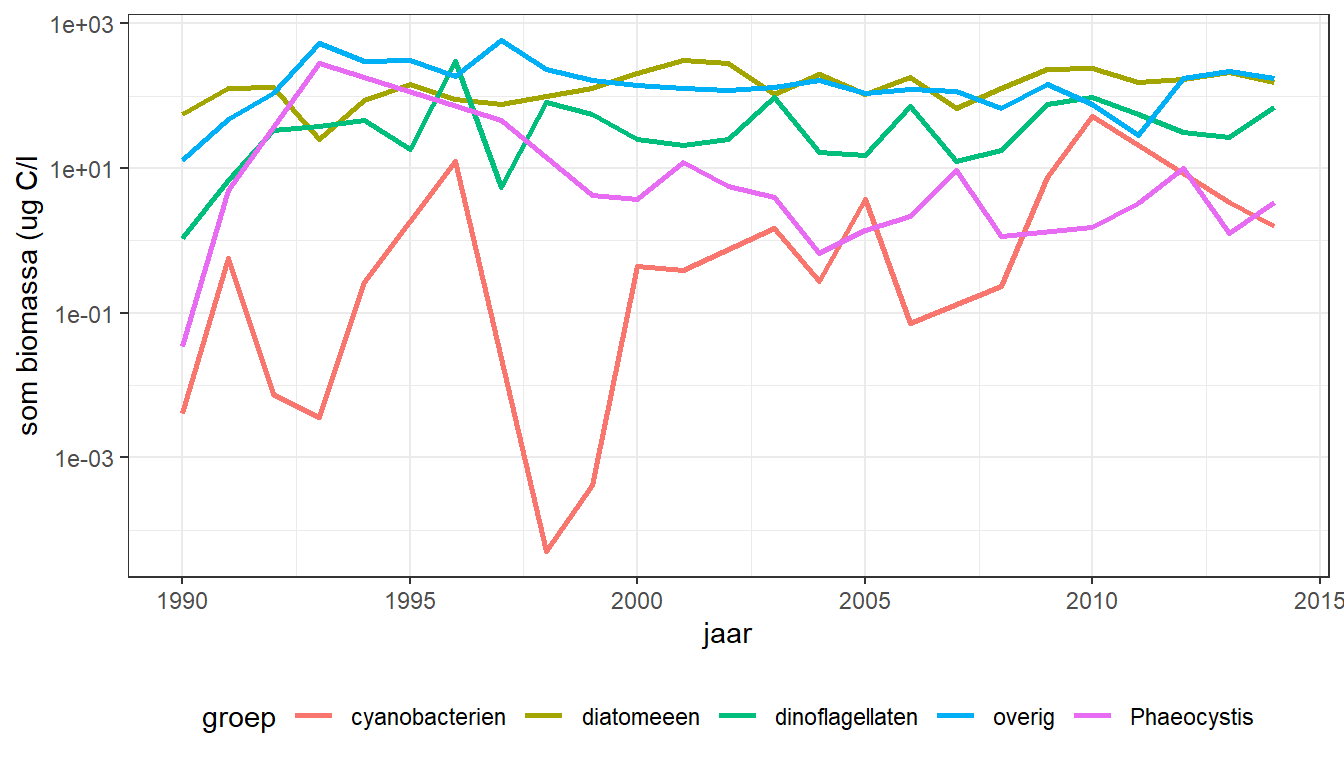
Figuur 9.1: Verloop van jaargemiddelde zomerbiomassa (logschaal!) voor fytoplanktongroepen in de tijd. In de jaren voor 2000 zijn geen geharmoniseerde metingen beschikbaar.
De biomassa (in µgC/l) van fytoplankton per groep is in figuur 9.2 zo uitgezet dat ze in relatie tot de totale biomassa gezien worden. Dit is alleen voor de periode 2006 - 2016 hier gedaan. Diatomeeën, samen met overige soorten, nemen het grootte deel van de biomassa voor hun rekening. In 2013 en 2014 worden zeer hoge diatomeeënbiomassa’s gezien. Dinoflagellatern nemen in sommige jaren gedurende korte tijd in de zomer een aanzienlijk deel in van de totale biomassa.
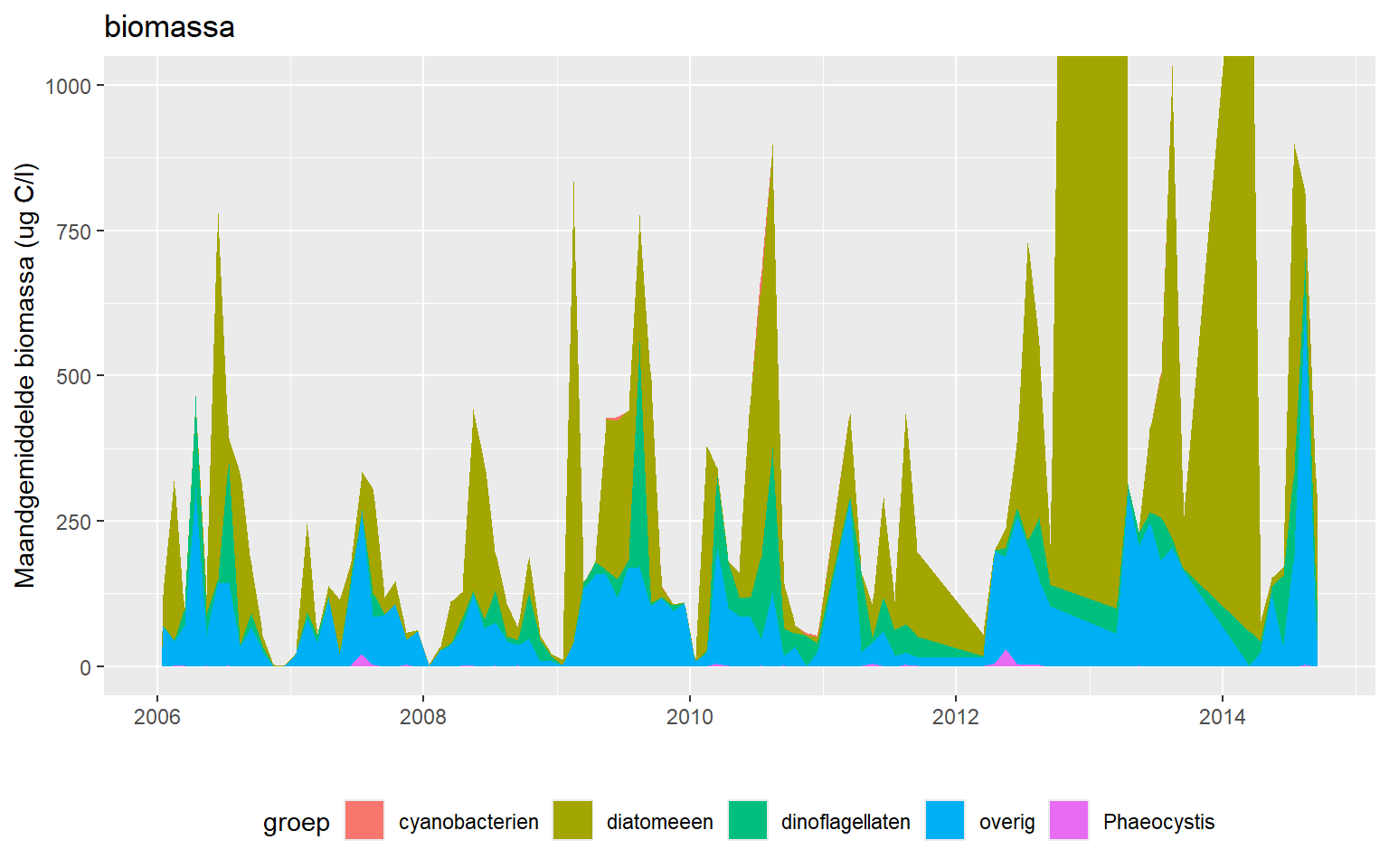
Figuur 9.2: Biomassa van de verschillende fytoplanktongroepen in het Grevelingenmeer over de periode 2006 - 2016.
Figuur 9.3 geeft het verloop van aantal cellen van de vier indicatorgroepen van januari tot en met december weer in box-whisker plots; let op de logaritmische schaal van de y-as (ug C/l). De linkerplot betreft de concentratie in het water nabij de bodem, de middelste plot nabij de spronglaag, en de rechterplot in het oppervlaktewater. Alhoewel de vergelijkende analyse met licht, zwevend stof en nutriënten nog moet plaatsvinden, kunnen enkele observaties al worden gedaan die wijzen op een afwijkend verloop ten opzichte van kustwateren, en duiden op dito afwijkende aansturing door licht en nutriënten. Het verloop van de concentratie fytoplankton in het oppervlaktewater is van laag in de winter naar hoog in de zomer. Een voorjaarsbloei is zichtbaar bij de diatomeeën, die evenals in andere wateren relatief vroeg bloeien ten opzichte van andere soortsgroepen. In het Grevelingenmeer ligt de piek zelfs dan relatief vroeg ten opzichte van andere wateren zoals in de kustzone (piek autotrofen na Julian dag 100, d.w.z. april), vermoedelijk vanwege het relatief heldere water in het Grevelingenmeer. De overige groepen tonen deze vroege voorjaarsbloei niet (of zeer zwak), maar eerder een toename die doorloopt tot in de zomer (juni) en daarna langzaam afneemt. Overigens vertoont de diatomeeënbloei een tweede piek rond juni, die boven die in februari uitkomt. Deze afname na de februaribloei duidt op het limiterend worden van silicaten, terwijl de toename van diatomeeën daaropvolgend aangeeft dat dit niet het geval is. Daarnaast duidt de continue toename van algenbiomassa tijdens de zomer (lijkt op een verband met de hoeveelheid licht) niet op een tekort aan voedingsstoffen maar op een lichtbeperking. Gezien de aanwezigheid van een spronglaag is de continue hoge algenmbiomassa zeker opvallend; meestal leidt een spronglaag tot een tekort aan voedingsstoffen omdat deze worden ‘opgesloten’ onder de spronglaag.
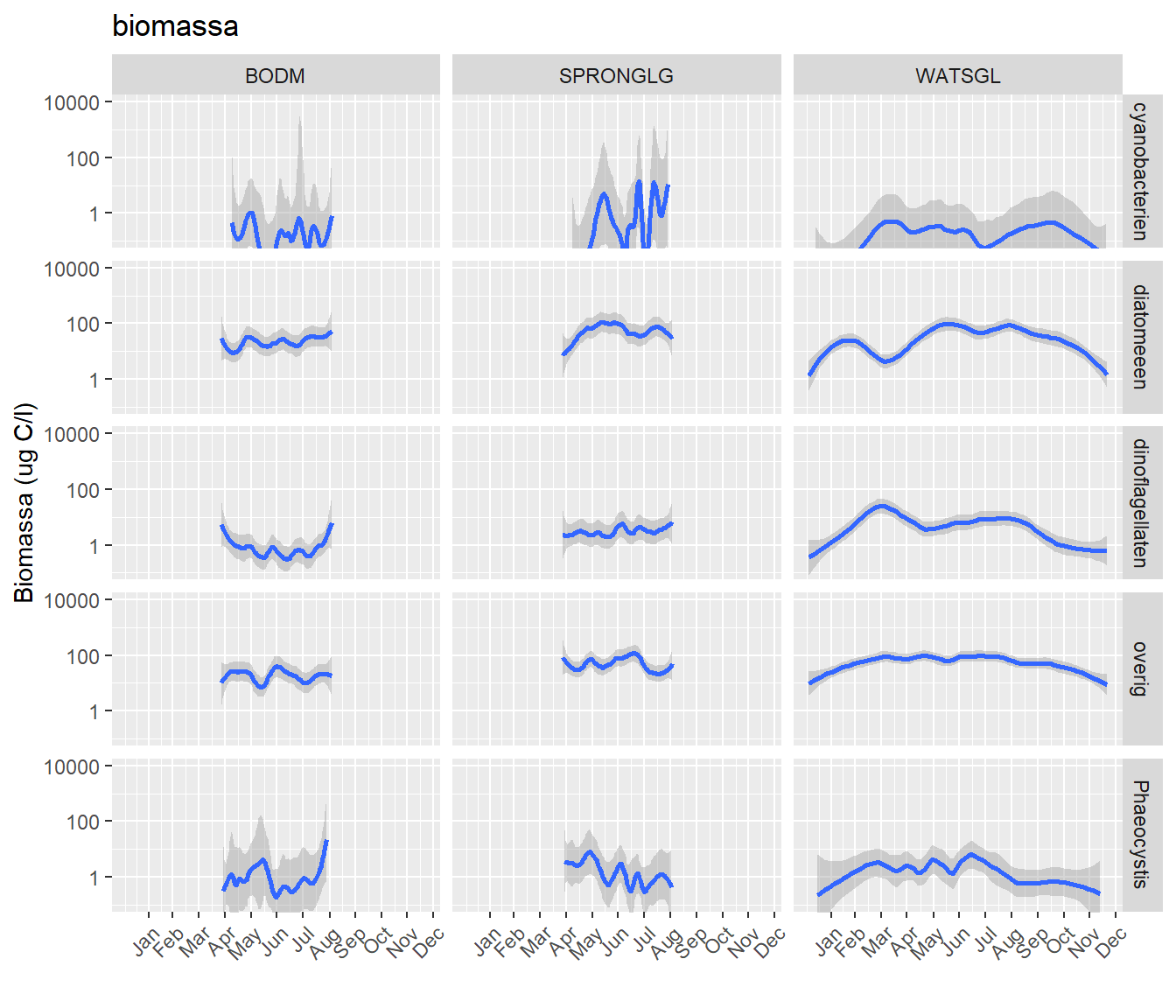
Figuur 9.3: Een samenstelling van alle fytoplanktondata (1990-2014) waarbij de verdeling van over het jaar is weergegeven in boxplots per week voor de verschillende groepen.
De concentraties van algen variëert van 10 ug C/l in de winter tot 100 ug C/l in de zomer voor diatomeeën en dinoflagellaten. Phaeocystis concentraties zijn veel lager: van 0,1 tot en met 10 ug C/l. Overige algenbiomassa varieert tussen de ca. 100 en 1000 ug/l.
De verhouding tussen de biomassa in oppervlaktemonsters enerzijds en de spronglaagmonsters anderzijds toont geen sterke duidelijke trend in de verdeling van algenbiomassa over de waterkolom tijdens de zomer (figuur 9.4); in de ratio oppervlakte/spronglaag is een lichte daling te zien gedurende de zomer voor de diatomeeën en de dinoflagellaten: dit duidt op een uitzakking van deze algen, hetgeen ook enigszins verwacht kan worden. In het najaar is er weer een stijging, wat betekent dat de algen voornamelijk aan het oppervlak voorkomen. De trend is dat de concentraties in de spronglaag en de oppervlaktemonsters veelal wat hoger zijn in de bodemmonsters (ratio lowess>1, maar de variatie is groot).
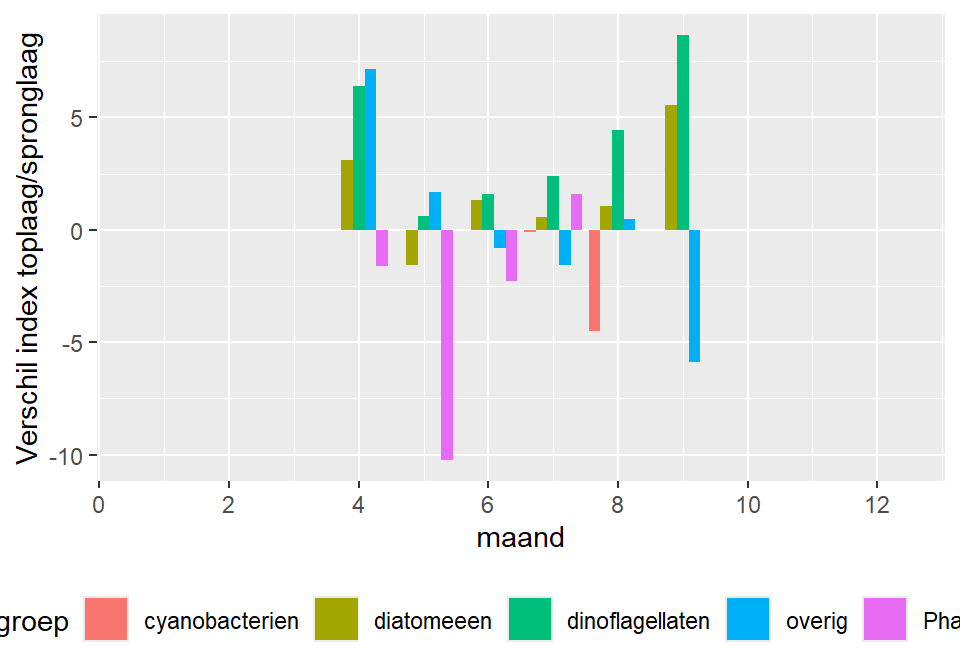
Figuur 9.4: Verhouding tussen fytoplankton biomassa aan de oppervlakte en in de spronglaag. Negatieve getallen geven aan dat de concentratie in de spronglaag hoger is, positieve getallen dat de concentratie aan het oppervlakte hoger is.
9.3 Schadelijke fytoplanktonsoorten
Er zijn een aantal phytoplanktonsoorten in de Grevelingen aangetroffen die bekend staan om hun schadelijke effecten. Een van de bekendste is Dinophysis acuminata, die door ophoping in mosslen diarrhetic shellfish poisoning (DSP) kan veroorzaken. Deze soort komt voor, en lijkt ook de laatste jaren in iets hogere aantallen voor te komen. De toxines van deze alg worden routinematig gemeten in productiemosselen in bijvoorbeeld Oosterschelde. Voor zover bekend zijn er tot nu toe nog geen mosselen gevonden met deze toxines.
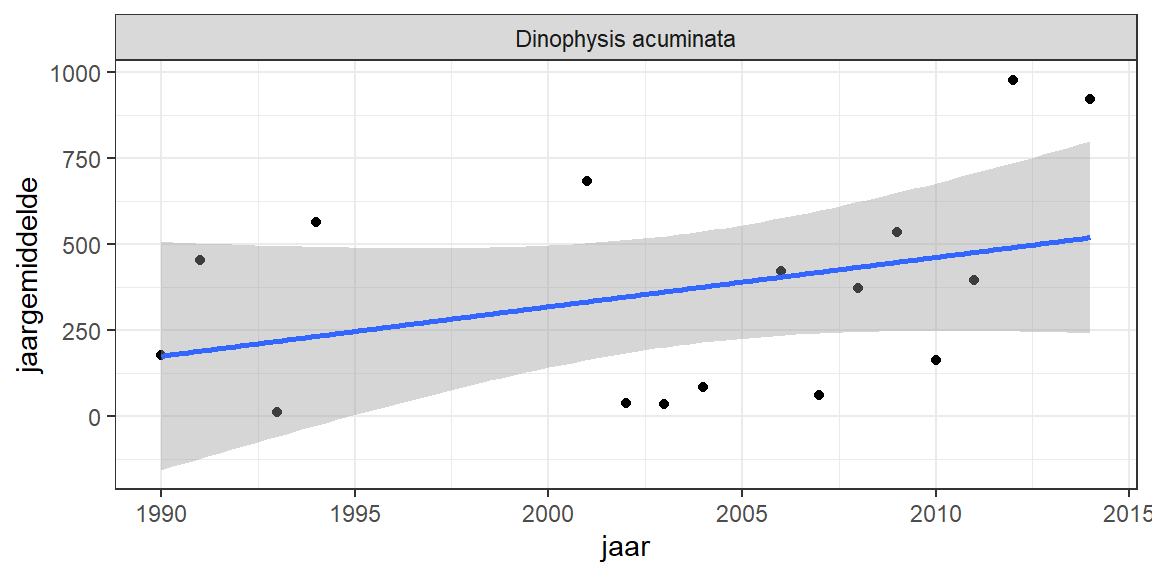
Figuur 9.5: Jaargemiddelde biomassa van Dinophysis acuminata, een alg die diarrhetic shellfish poisoning (DSP) kan veroorzaken.
9.4 Schadelijke fytoplanktonsoorten in kreken
In Zeeland zijn er in de afgelopen 20 jaar giftige dinoflagellatensoorten aangetroffen in verschillende kreken verspreid over de gehele provincie (Figuur 9.6). Het gaat daarbij vooral om soorten binnen de genera Alexandrium en Gymnodinium (beide zijn overigens lastig te onderscheiden), die het vooral goed doen in brak water (Martens et al. (2016)). Sinds 2012 is er een terugkerende bloei van Alexandrium ostenfeldii (Burson et al. 2014; Brandenburg et al. (2017)). De soort uit de Ouwerkerksekreek maakt verschillende neurotoxines aan, zoals saxitoxines, spirolides en gymnodimines (Van de Waal et al. (2015)). Het is zeer aannemelijk dat er in veel meer kreken dergelijke giftige algenbloeien jaarlijks terugkeren, gezien ook de waarnemingen in het verleden. Een bloei met hoge celaantallen gaat samen met verhoogde concentraties toxines, die een direct risico vormen voor eventuele recreanten maar ook voor dieren, met name watervogels en mogelijk ook huisdieren. Daarnaast kunnen de toxines ophopen in aangrenzende mosselpercelen als de kreken daarop spuien. Desondanks is er momenteel geen monitoringsprogramma om dit risico in kaart te brengen.

Figuur 9.6: Verspreiding giftige algen in kreken in Zeeland. Een indicatie geeft aan dat er een mogelijk giftige soort in de kreek is waargenomen in de afgelopen 20 jaar. Gegevens uit nlbif, limnodata.
9.5 Benthische micro-algen
Er is slechts eenmalig en op een station één opname geweest van benthische microalgen (microfytobenthos) voor zover nu bekend. Op grond hiervan is niets te zeggen over de populatie microfytobenthos, verspreiding, of trends.
9.6 Macro-algen
Dit deel wordt later nog verder ingevuld. Voor het moment wordt verwezen naar de vorige rapportage en de delta-expertise wiki
9.7 Zoöplankton
Er is zeer weinig bekend over de status en ontwikkeling van zoöplankton. Het zoöplankton vertegenwoordigt een diverse groep met als belangrijkste diergroepen kreeftachtigen zoals eenoogkreeftjes, (rib)kwallen, maar ook larven van o.a. vissen, grotere kreeften en krabben, zeesterren, zee-egels, wormen en schelpdieren. Zoöplankton is de schakel tussen algen (de primaire producenten) en planktivore vis, die op zijn beurt weer ten prooi valt aan roofvis en visetende vogels. De potentiële graasdruk van zoöplankton is een belangrijke parameter om het ecologisch functioneren van watersystemen te beschrijven (STOWA, 2010). Toch is deze groep geen ‘kwaliteitselement’ in de Europese Kaderrichtlijn Water en wordt daarom ook niet gemonitord in het Grevelingenmeer. Van het Grevelingenmeer is bekend dat er enorme bloei van de ribkwal Mnemiopsis leidyi maar ook van oorkwallen kan optreden die ongetwijfeld een substantiële invloed zullen hebben op het voedselweb door o.a. consumptie van algen en ander zoöplankton (inclusief schelpdier- en vissenlarven).
9.8 Benthische fauna inclusief schelpdierbestanden en hard substraat biota
De analyses voor macrofauna richten zich op het macrobenthos. De term ‘macrofauna’ heeft betrekking op alle ongewervelde dieren in het Grevelingenmeer. De ‘macrobenthische fauna’ of ‘macrobenthos’ betreft de ongewervelde dieren die in, op of vlak boven de bodem leven.
Methode
De ontwikkelingen van het macrobenthos in het Grevelingenmeer zijn deels bepaald aan de hand van data uit het MWTL-programma en deels gebaseerd op de literatuur. Als onderdeel van het MTWL-programma vindt er sinds 1990 een inventarisatie plaats van het macrobenthos van het Grevelingenmeer met als doel inzicht te krijgen in de ruimtelijke en temporele variatie van de benthische fauna. De resultaten van deze meetcampagnes zijn verwerkt in diverse rapporten met het meest recentelijke overzicht in Verduin et al. (2018). Daarnaast heeft recentelijk een uitgebreide analyse plaatsgevonden in opdracht van het ministerie van LNV en Staatsbosbeheer welke de kwantitatieve ontwikkelingen beschrijft van het macrobenthos in de periode 1992-2016, gebruik makend van de MTWL-data (Mulder et al. 2019). Jaartallen 1990 en 1991 zijn niet meegenomen in deze analyse omdat de data uit deze twee jaren niet gewaarborgd kon worden (onbetrouwbare data). In Mulder et al (2019) wordt ook een overzicht gegeven van de frequentie van bemonsteringen en de verschillende bemonsteringsmethodes die zijn gebruikt in deze periode. Door middel van literatuuronderzoek (o.a. Nienhuis 1978, Lambeck 1986, Lambeck & Pouwer 1986, Lambeck & Smet 1987, Fortuin & Altena 1990) is tevens een overzicht gemaakt van de ontwikkelingen in de periode voor 1990.
Lambeck (1986) geeft een duidelijk overzicht van de ontwikkelingen na de afsluiting van het Grevelingenmeer. De beschrijving hieronder baseert zich voornamelijk op drie eerdere studies: Lambeck (1982), Wolff et al. (1977) en Wolff & De Wolf (1977). Voor de afsluiting in mei 1971 is uitgebreid onderzoek gedaan naar de biomassa en productie van het macrobenthos. Dit onderzoek diende als een referentie voor toekomstige veranderingen na de afsluiting (Wolf & De Wolf 1977). Direct na de afsluiting is een regulier bemonsteringsprogramma voor macrobenthos opgezet waarbij jaarlijks in het voorjaar bemonsterd werd. De eerste fase van dit programma liep tot 1977 (Wolf et al. 1977, Lambeck 1982) met als doel onderzoek te doen aan macrobenthos in het Grevelingenmeer. Vanaf 1981 ging de tweede fase van start waarbij het verkrijgen van referentiemateriaal voorop stond. In deze periode is vrijwel jaarlijks een bestandsopname uitgevoerd (o.a. Lambeck & Pouwer 1986, Lambeck & Smet 1987, Fortuin & Altena 1990). Deze gegevens dienden als referentiemateriaal voor studies aan andere componenten van het systeem. Daarnaast keek het onderzoek naar de effecten van de net geopende spuisluis in de Brouwersdam en gaf het inzicht in lange-termijn trends van belangrijke en dominante soorten. Verdere details over bemonsteringslocaties en technieken zijn te vinden in de bovengenoemde rapporten en publicaties.
Als deel van het MWTL-programma vindt er sinds 1990 in het voor- en najaar een bemonstering plaats in het oostelijk en westelijk deel van het Grevelingenmeer (Figuur 9.7). In beide deelgebieden worden monsterlocaties evenredig verdeeld over drie diepte strata: 0-2 m, 2-6 m en dieper dan 6 m (t.o.v. NAP = zomerpeil + 0.20 m vóór 2004, zomerpeil + 0.26 m na 2004) (Verduin et al. 2018). Sinds 1994 wordt in ieder deelgebied en binnen elk dieptestratum op vastgestelde locaties bemonsterd. Dit resulteert in 120 monsters per jaar, 60 in het voorjaar en 60 in het najaar. Na 2013 is de jaarlijkse macrozoöbenthosmonitoring van het MWTL programma teruggeschroefd naar 1 x per drie jaar, wel nog in het voor- en najaar. Verdere details zijn te vinden in Verduin et al. (2018) en Mulder et al. (2019).

Figuur 9.7: Ligging van de macrobenthos bemonsteringspunten (rood) in het oostelijk en westelijk deel van het Grevelingenmeer in de periode 1992 tot 2016 (MWTL).
Resultaten
Aan de hand van literatuuronderzoek en data uit het MWTL-programma wordt een overzicht gegeven van de ontwikkelingen van biomassa (g/m2) van het macrobenthos in de periode na de afsluiting van de Grevelingen tot 2016 (Figuur 9.8). De biomassa in Figuur 9.8 is een gewogen jaargemiddelde wat betekent dat de biomassa is gecorrigeerd voor het bemonsteringsoppervlak in de verschillende dieptestrata waardoor de werkelijke biomassa beter wordt weergegeven. Figuur 9.8 is samengesteld uit drie verschillende datasets: Lambeck (1986), bemonsteringen door het Delta Instituut voor Hydrobiologisch Onderzoek (DIHO, nu NIOZ Yerseke) en MWTL-data. De biomassa nam sterk af in de eerste jaren na de sluiting (Figuur 9.8, Lambeck, 1986) maar herstelde zich en piekte tussen 1995 en 2000 waarna de biomassa weer gestaag afnam. Hieronder worden deze ontwikkelingen verder besproken.

Figuur 9.8: De ontwikkeling van de gewogen (gecorrigeerd voor bemonsteringsoppervlak in dieptestrata) gemiddelde biomassa (gram per m2) van bodemdieren macrobenthos in het Grevelingenmeer in de periode 1971-2016, samengesteld uit drie verschillende datasets/bronnen: Lambeck 1986 (blauw), DIHO-bemonsteringen (groen) en MWTL-data (paars).
Periode 1971-1978 na de afsluiting
Na de afsluiting is het Grevelingenmeer armer geworden in het soortenaantal. De daling bedroeg ruim een kwart: van ca. 338 soorten in 1964-1970 naar ca. 252-264 soorten in 1972 (Nienhuis 1978, Lambeck 1986). Nieuwe en zeer algemene soorten waren de gevlochten fuikhoorn en de brakwaterkokkel. Lambeck (1986) gaf aan dat ook de platte oester als nieuwe soort gezien zou kunnen worden. Het bestand was na de strenge winter van 1962/63 bijna geheel verdwenen maar heeft zich na 1971 sterk uitgebreid. Een grote daling deed zich voor bij de krabben en kreeften. Van de oorspronkelijk negen soorten hebben alleen de strandkrab en de heremietkreeft zich weten te handhaven. Ook verdwenen alle zeespinnen en bleef onder de stekelhuidigen slechts de zeester over (Lambeck 1986).
Onder de soorten die zich hadden weten te handhaven vonden forse verschuivingen plaats. Daarnaast kregen deze soorten gezelschap van soorten die lage stroomsnelheden tolereerden. Zo werd de gevlochten fuikhoorn voor het eerst aangetroffen in 1973 en kwam in 1976/77 in het hele meer voor met dichtheden van 160 ind/m2 in zones dieper dan 1.5 m (Lambeck 1986). Een andere belangrijke soort was de mossel welke een brede verspreiding aan zoutgehaltes kan tolereren en een enorme voortplantingscapaciteit heeft. Na een terugval in 1971 leek het weer beter te gaan met de mossel en was 1976 voor vele soorten een zeer goed jaar met een ruime verdubbeling van het macrobenthosbestand wat voornamelijk te danken was aan de mossel (Figuur 9.8, Lambeck 1986). Lambeck geeft als reden voor de enorme toename van het bestand aan dat op de koppen van de platen en in de diepere geulen, zo’n 20% van de open Grevelingen, de stroming en sedimentbewegingen vroeger zo heftig waren dat alleen specialisten zich daar konden vestigen. Na het wegvallen van het getij konden deze gebieden gekoloniseerd worden (Lambeck 1986). Daarnaast suggereert hij dat de afname van grote predatoren zoals vissen en krabben ook voordelig zou zijn geweest, naast het wegvallen van de predatie door steltlopers bij laagwater. Echter, door zo’n enorme toename schoot het systeem door zijn plafond en al snel was er niet genoeg voedsel beschikbaar.
De mosselen in het Grevelingenmeer werden minder groot en vooral minder zwaar. Vooral 1977 was een zwaar jaar voor mosselen. Het duurde tot 1981 voordat de oude mosselen weer konden groeien en zich voorzichtig aan weer konden gaan voorplanten, maar pas in 1983 was sprake van een geslaagde voorplanting (Lambeck 1986).
Periode 1979-2016 na opening van de Brouwerssluis
Even daarna vond nog een systematische verandering plaats, waarbij de in december 1978 geopende Brouwerssluis een belangrijke rol speelde. Het inlaten van zout Noordzeewater leidde in het eerste jaar tot stratificatie waardoor vrijwel alle macrobenthos beneden de 8 m zou zijn doodgegaan (Lambeck 1986). Een volgende ontwikkeling die plaatsvond, was het ophopen van organisch materiaal in de diepere delen van het meer door het wegvallen van het getij. Doordat al dit materiaal werd afgebroken steeg de zuurstofvraag en verslechterde hierdoor de zuurstofomstandigheden nabij de bodem. Daarnaast raakten schelpen en andere harde ondergronden ondergesneeuwd door het organisch materiaal waardoor broedval steeds moeilijker werd (Lambeck 1986). Om deze reden vertoonde de oester in de jaren 1981-1984 een duidelijke verschuiving naar ondieper water. Terwijl dit eerst alleen van belang was voor een relatief beperkt oppervlak in de diepere delen, verschoof de grens snel op naar 7 m diepte, waardoor bijna 1/5 van het meer verarmde. De periode 1981-1984 wordt dan ook gekenmerkt door een lage biomassa (Lambeck 1986).
Vanaf 1980 werd de sluis alleen in de wintermaanden gebruikt en bleef het zoutgehalte zo op een stabiel hoog niveau. Met het Noordzeewater zijn allerlei verdwenen soorten in het Grevelingenmeer teruggekeerd (Lambeck 1986). De voedselsituatie heeft zich ook verbeterd in de jaren tachtig met een forse stijging van de hoeveelheid algen. Het dalen van de dichtheid en gewichtshoeveelheid van mosselen bood een kans voor andere soorten om dit gat op te vullen. Zo nam het muiltje in deze periode flink toe en bereikte in 1986 en 1988 dichtheden van enkele honderden exemplaren per vierkante meter in het gebied van 2 tot 6 m diepte (Lambeck & Smet 1987). In 1989 is de verdeling van de biomassa over de dieptestrata sterk veranderd waarbij de gemiddelde biomassa in het diepste stratum (>13.5 m) verdrievoudigd is in vergelijking met 1988. Hierdoor is er geen verschil meer in biomassa tussen de verschillende strata. Vooral soorten zoals het muiltje, mossel, oester en zeepokken komen nu met een hoge biomassa voor in het diepste stratum (Fortuin & Altena 1990).
Tussen 1990 en 2000 treden er geen grote veranderingen op en blijven de totale biomassa en aantallen relatief gelijk (Figuur 9.9) waarbij aantallen werden gedomineerd door wormen, weekdieren en geleedpotigen, en biomassa door filtrerende weekdieren (Schaub et al. 2002, Mulder et al. 2019). Wel was de gemiddelde biomassa vrij hoog (70 g/m2). De biomassa was hoger dan destijds in de Oosterschelde en Westerschelde werd gevonden en iets lager dan in het Veerse Meer.

Figuur 9.9: De gemiddelde totale biomassa (gram per m2, links) en dichtheid (aantal per m2, rechts) per jaar in de periode 1992 tot 2016. Het grijze gebied geeft de standaard error weer.

Figuur 9.10: Gemiddelde biomassa (gram per m2) per jaar van Mytilus edulis (mossel, links), Crepidula fornicata (muiltje, midden) en Magallana gigas (ook wel Crassostrea gigas, Japanse oester, rechts). Het grijze gebied geeft de onzekerheid uitgedrukt in standard error van het gemiddelde weer.
Na 2000 lijkt het aandeel van het muiltje in de totale biomassa echter flink af te nemen van 64% in 1993 naar 29% in 2010 (Figuur 9.10). De plaats van het muiltje wordt ingenomen door de Japanse oester die vanaf 2000 langzaam in aantallen en biomassa toeneemt (aandeel: 0% in 1993 en 31% in 2010, Figuur 9.10) (Mulder et al. 2019). Echter, de totale biomassa is sterk afgenomen met 88 g/m2 in 1993 naar ongeveer 25 g/m2 in 2009 (Figuur 9.9). Het lijkt er wel op dat de biomassa weer wat toegenomen is in 2010 maar weer afneemt in 2013 en 2016. Gedurende deze periode is de soortensamenstelling van de macrobenthosgemeenschap ook nog steeds aan het veranderen, met een structureel verschil tussen de ondiepe (0-2 m) en diepere delen (>2 m) (Figuur 9.11)(Mulder et al. 2019). Wanneer de samenstelling van de macrobenthosgemeenschap gelijk is tussen twee jaren dan vallen de punten (jaren) in Figuur 9.11) op elkaar zoals het geval is in 2006 en 2007 in de diepte strata <2 m. Verschilt de samenstelling echter tussen jaren dan worden de punten in deze analyse verder uit elkaar gezet (bijv. 2008 en 2009). Hoe groter de afstand tussen de punten, hoe groter het verschil is in de samenstelling van de macrobenthosgemeenschappen. De samenstelling is bijvoorbeeld veranderd in 2000 ten opzichte van de periode 1992-1999 waar de punten vrij dicht op elkaar liggen (Figuur 9.11)). Daarnaast laat Figuur 9.11) zien dat dat de samenstelling van de macrobenthos in de ondiepe zone (0-2 m) verschilt van dat van de twee diepere strata waar de samenstelling meer overeenkomt. Dit laat zien dat bijna vijftig jaar na de afsluiting van het Grevelingenmeer, de bodemdiergemeenschap zich nog steeds ontwikkelt in alle strata en de ontwikkeling ook plaatsvindt in dezelfde richting. Het is niet zo dat gemeenschappen zich verschillend ontwikkelen op bepaalde dieptes.

Figuur 9.11: nMDS plot met weergave van jaar-centroïden binnen elke dieptestratum. Opeenvolgende jaren (tussen 1992 en 2016) zijn verbonden met pijlen, uit Mulder et al. (2019)
De ruwe data (Figuur 9.12) en de toegepaste modellen in Mulder et al. (2019) geven aan dat waterdiepte een significant effect heeft op biomassa en dichtheid (p<0.001). In de ondiepe vooroever is de biomassa laag maar neemt significant toe tot een diepte van ongeveer 6 m. Vanaf 6 m onder NAP neemt de biomassa weer significant toe tot ongeveer 15 m waar de betrouwbaarheid van de gegevens afneemt door het geringe aantal monsters dat genomen is op deze diepte. De relatie tussen waterdiepte en biomassa (of dichtheid) blijft hetzelfde voor alle dominante taxonomische groepen zoals Mollusca, Annelida en Arthropoda (Mulder et al. 2019).

Figuur 9.12: De verhouding van de gemiddelde biomassa (gram per m2) over de diepte (m t.o.v. NAP) in de periode 1992 tot 2016.
De studieperiode is ook opgedeeld in twee (voor- en na 1999) en vier perioden (5-6 jaar intervallen) waarna gekeken is of deze relatie veranderde of verschoof over de tijd. De analyses zijn niet opgenomen in het rapport door Mulder et al. (2019) maar de resultaten laten zien dat de optimale biomassa blijft liggen tussen de 4-6 m onder NAP met een significante daling naar diepere en ondiepere delen zoals ook waarneembaar in Figuur 9.12.
Verhouding wormen/schelpdieren
Schaub et al. (2002) suggereren dat het Grevelingenmeer minder gunstig is geworden voor soorten die voedsel uit het water filtreren. Door de afsluiting van het gebied zakt zwevend materiaal eerder naar de bodem waarvan bodemetende organismen kunnen profiteren (Schaub et al. 2002). Vanaf 1990 blijkt de verhouding van de wormen ten opzichte van de weekdieren toe te nemen waardoor er sprake zou zijn van een zogenaamde ‘verworming’ van het systeem (Schaub et al. 2002). Het voortzetten van die trend werd gesignaleerd door Wetsteijn (2011) tot 2008 en bevestigd door de huidige aanvulling van de MWTL-monitoringsreeks tot 2016. Door dezelfde berekening uit te voeren als in voorgaande rapporten blijkt dat de verhouding van de wormen ten opzichte van de weekdieren nog steeds significant toeneemt (y=0.922 x – 1767, p<0.001, R2=0.44, Figuur 9.13). Uitgedrukt in biomassa wordt voor deze periode geen significante trend gevonden (y=0.231x – 453, p=0.09, R2=0.05). Echter, de term ‘verworming’ is onjuist. Verhoudingsgewijs neemt het relatieve aandeel wormen wel toe, maar dat komt omdat het relatieve aandeel schelpdieren afneemt. De afname van schelpdieren is zelfs te herleiden tot de afname van één soort: het muiltje (Mulder et al. 2019).

Figuur 9.13: Verworming in het Grevelingenmeer in de periode 1992-2016 uitgedrukt in de ratio worm/(worm+weekdier) dichtheid voor de periode 1992-2016 met gegevens in het oostelijk en westelijk deel samengenomen.
Samenvattend
Het macrobenthos onderging grote wijzigingen in de tijd, mede door veranderingen in het sluisbeheer. Na de afsluiting is het Grevelingenmeer armer geworden in soortenaantal. Het inlaten van zout Noordzeewater door de opening van de Brouwerssluis leidde in het eerste jaar tot stratificatie waardoor vrijwel alle macrobenthos beneden de 8 m zou zijn doodgegaan. Een volgende ontwikkeling die plaatsvond, was het ophopen van organisch materiaal in de diepere delen van het meer door het wegvallen van het getij, waardoor de leefomstandigheden voor het macrobenthos hier verslechterde. De biomassa van het macrobenthos in het Grevelingenmeer wordt gedomineerd door filtrerende schelpdieren, maar de dominante soorten veranderden in de loop der tijd. Sinds 1990 wordt de biomassa in belangrijke mate bepaald door exoten: het muiltje en de laatste jaren de Japanse oester. De benthosgemeenschappen zijn in de periode 1992 – 2016 veranderd, en dit in zowel de diepe als ondiepe zones van het meer. Dit suggereert dat de macrobenthos gemeenschappen 50 jaar na afsluiten nog steeds in ontwikkeling zijn in reactie op het afsluiten van het meer en het gevoerde waterbeheer en niet alleen beïnvloed worden door het optreden van zuurstofloosheid in het meer. Er is sprake van een systeem brede ontwikkeling waarbij op alle diepten (dus ook in de ondiepe zone) veranderingen optreden.
9.9 Zeegras
Introductie: In het Grevelingenmeer bereikte Groot zeegras een maximale bedekking van 4600 ha in 1978 (Nienhuis et al 1996). Na veranderingen van het waterbeheer begin negentiger jaren is het zeegras in het Grevelingenmeer snel geslonken tot minder dan 100 ha in 1993 en was enkele jaren later geheel verdwenen. Aanvankelijk dacht men aan een Labyrinthula schimmelinfectie (NRC 1991), invloed van stress op een verarmde populatie (Nienhuis et al 1996) en ongunstige siliciumconcentraties (Herman et al 1996). Uit proeven bleek dat zeegrassen (uit het Grevelingenmeer) geen invloed ondervonden van lagere siliciumgehaltes (Kamermans et al. 1999), maar wel langzamer groeiden bij hogere zoutgehaltes (Kamermans et al. 1999, van Katwijk et al. 1999). Toch zijn zeegrassen over het algemeen aangepast aan zoutgehaltes tot 34 promille of hoger (Giesen et al 1990). Ook is het milieu in het Grevelingenmeer in alle andere opzichten heel geschikt voor zeegras, zo bleek uit een vergelijkende studie naar de water en bodemkwaliteit van 84 NW-Europese wateren (van der Heide et al. 2009). Een aannemelijke verklaring voor het verdwijnen in de jaren 90 is daarom dat de zeegraspopulatie in de jaren 70 bij het sterk dalende zoutgehalte (tot 14-16‰) is uitgeselecteerd op lage zoutgehaltes. Toen het zoutgehalte in het Grevelingenmeer weer steeg van 24‰ naar meer dan 30‰ stierf het aan laag-zoutgehalte aangepast zeegras uit; door een genetische bottleneck was het niet meer in staat de omslag te maken. Kiemingsexperimenten ondersteunen dit: Grevelingenplanten konden niet meer kiemen bij hoge zoutgehaltes (van Katwijk, ongepubliceerde resultaten van proeven 1989-90). Inmiddels is vastgesteld dat de omstandigheden op een aantal locaties op dit moment, onder meer door het voeren van een natuurlijker waterbeheer en lager zoutgehalte, weer kansrijk zijn voor groot zeegras. Rijkswaterstaat voert i.s.m. andere partijen experimenten uit om mogelijkheden voor herstel van zeegras in het Grevelingenmeer nader te onderzoeken. Bron: RWS in WMR rapport C089/19 paragraaf 1.4.2.
Uitwerking (RWS): Groot zeegras was ooit algemeen in het Grevelingenmeer en Veerse Meer maar is de afgelopen twee decennia verdwenen door veranderingen in zoutgehalte (Grevelingenmeer) of ten gevolge van eutrofiëring (Veerse Meer). Nu lijken de milieuomstandigheden weer gunstig voor Groot zeegras, en in het kader van de Kaderrichtlijn Water (KRW) wordt een pilot uitgevoerd waarin onderzocht wordt of Groot zeegras kan worden teruggebracht in deze Zeeuwse wateren. Inmiddels is een eerste “proof of principle” fase uitgevoerd in 2016-2017 waarbij zaden zijn verspreid in het Grevelingenmeer volgens een aantal methoden, en een eerste monitoring wijst op een succesvolle kieming.
Zeegrasherstelprojecten in Zeeland zijn om verschillende redenen van belang: i) zeegrasvelden kunnen golven beïnvloeden en sediment vasthouden en dragen bij aan kustbescherming; ii) ze vormen kraamkamers voor de visserij en zijn van ecologisch en economisch belang; en iii) Nederland heeft een inspanningsverplichting aangezien zeegrasvelden in Europees verband een beschermd habitat vormen.
In de 2e tranche van de KRW heeft Rijkswaterstaat Zee- en Delta bij wijze van herstelmaatregel proeven met zeegras in de beide zoute meren in Zuidwest Nederland opgevoerd, ten einde dit te herintroduceren. Rijkswaterstaat voert in 2017-2021 een meerjarige pilot uit die rekening houdt met de lessons learned van de uitzaaiproeven die hebben plaatsgevonden in Grevelingenmeer (2014 en 2016-2017) en Waddenzee (2003 - doorlopend). Rijkswaterstaat wil een grotere kans van slagen bereiken door vooraf ook de mogelijke risico’s in kaart brengen.
Groot zeegras vormt een belangrijke soort en zeegrasvelden vormden ooit een belangrijk habitattype in het zuidwestelijk Deltagebied. Omdat het zeegras in dit gebied geen directe economische betekenis had zoals in het Waddengebied werd er tot de vijftiger jaren weinig aandacht aan besteed. Na de afsluiting van de Zandkreek (1960), Veerse Gat (1961) en Grevelingen (1971) breidde Groot zeegras zich sterk uit in de nieuw gevormde brakke meren.
Wereldwijd komen 63 soorten zeegras voor, en in Europa circa 6 soorten. In Nederland komen 3 soorten zeegras voor, Zostera marina, Z. noltii en Ruppia maritima. In dit project focussen we op Z. marina, groot zeegras, dat voorkwam in het Grevelingenmeer en het Veerse Meer. In de Oosterschelde komt ook Z. marina voor, maar in veel mindere mate. Het leek daar bijna verdwenen, maar onlangs is een nieuwe populatie opgekomen op de slikken van Oude Tonge. (In de Oosterschelde komt vooral Z. noltii voor.) Groot zeegras kent twee vormen of types, een doorgaans eenjarig type met flexibele planten rond NAP, en een ondergedoken robuuster type die meerjarig is. De laatste is in de jaren 1930 uitgestorven in de Waddenzee, en heeft in Zeeland waarschijnlijk nooit gegroeid. In de Grevelingen is iets merkwaardigs gebeurd; de eenjarige vorm die er vóór afsluiting groeide rond NAP, heeft zich na afsluiting ontwikkeld tot een overwegend meerjarige vorm, maar wel van het flexibele type (b.v. Nienhuis et al. 1996).
Zeegras wordt wereldwijd gewaardeerd om zijn kraamkamerfunctie en habitatfunctie voor jonge vis, krabben en garnalen en voedselruif en schuilplaats voor grotere vissoorten. Minstens zo belangrijk is zijn ecosysteem engineering capaciteit (ook wel biobouwercapaciteit genoemd). Dit laatste bestaat op de eerste plaats uit het remmen van golfslag en stroming en daardoor stabiliseren van het sediment. Hierdoor ontstaan meerdere ecosysteemfuncties: het water wordt helderder, de bodem erodeert minder of hoogt op, en als gevolg van dat laatste helpt een zeegrasveld mee aan het ondiep houden van de vooroever van schorren en dijken (kustbeschermingsfunctie), en koolstof wordt opgeslagen (Orth et al. 2006, van der Heide et al. 2007, Fourqurean et al. 2012, Christianen et al. 2013, Mtwana Nordlund et al. 2016, Unsworth & Cullen-Unsworth 2017). Dit laatste is belangrijk om klimaatsverandering te mitigeren. De opslag van koolstof in de zee en in kwelders wordt blue carbon genoemd; zeegras draagt wereldwijd ongeveer 11% bij aan blue carbon (Nelleman et al. 2009). In het algemeen geldt dat hoe meer zeegras (meer hectare, meer dichtheid) hoe sterker de ecosysteem functies (Unsworth & Cullen-Unsworth 2017). Een ander gevolg van zeegrasaanwezigheid is een waterzuivering, niet alleen worden voedingstoffen (N en P) opgeslagen in het sediment net als koolstof, maar ook worden stikstofverbindingen versneld omgezet in stikstofgas, omdat denitrificatie-processen in de bodem versterkt worden door de zeegraswortels (b.v. Reynolds et al. 2016). Deze waterzuivering kan belangrijk zijn om algenbloei en anoxia te helpen voorkomen.
Wereldwijd wordt zeegras bedreigd. Net als in Nederland nemen de arealen ook elders in de wereld af, uitzonderingen daargelaten (Waycott et al. 2009). Dit heeft geleid tot een groot aantal aanplantprojecten. Wereldwijd is het succespercentage ongeveer 37%. Analyse van de wereldwijde aanplantprojecten leert dat hoe groter de schaal van aanplant, hoe groter de het succespercentage (N.B. dit is een gemiddelde, er zijn natuurlijk ook uitzonderingen) (van Katwijk et al. 2016). Dit wordt verklaard door twee fenomenen. Op de eerste plaats (i) is er een kritische massa nodig voor vestiging, omdat zeegras zijn eigen milieu verbetert. Een aantal losse planten zal bijvoorbeeld makkelijker wegspoelen dan een dichte patch. Zo zijn er nog veel meer voorbeelden van positieve feedbacks in zeegrasvelden (een overzicht hiervan wordt gegeven in Maxwell et al. (2016), die allemaal bijdragen aan het belang van het opbouwen van een kritische massa. Dit gebeurt op meerdere schalen; de sedimentstabilisatie speelt bijvoorbeeld een lokale rol voor niet-wegspoelen van de planten, maar op zeegrasveldschaal speelt sedimentstabilisatie een rol voor het helder maken van het water en kan op die manier het zeegras faciliteren. Op de tweede plaats (ii) is er grote variabiliteit in aanplantsucces, logischerwijs gerelateerd aan de grote natuurlijke omgevingsvariabiliteit. Omgevingsvariabiliteit is meestal niet te voorspellen (getijden natuurlijk wel); bijvoorbeeld een koud of warm voorjaar, of stormactiviteit. Temperatuur heeft geen belangrijk effect op het zeegras zelf, maar heeft effect op de mate van uitdroging van de bodem, op de activiteit van fauna (timing van wadpier en krabbenaanwezigheid), op bodembacteriën (meer of minder toxisch sulfide) etc.. Storm kan natuurlijk een zeegrasveld doen wegspoelen, als deze sterk genoeg is en uit de “verkeerde” hoek komt. IJsgang blijkt overigens doorgaans geen probleem te zijn voor zeegras in onze contreien. Het gevolg van de natuurlijke omgevingsvariabiliteit voor zeegrasaanplanten is dat een aanplant wisselend succes heeft, bijvoorbeeld in jaar 1 niet aanslaat, terwijl een zelfde aanplant in jaar 2 een groot succes kan zijn (zelfde locatie, zelfde omvang, zelfde donor etc). Ook is er vaak een grote ruimtelijke variabiliteit in succes: sommige aangeplante patches breiden zich uit, terwijl naastgelegen of tussengelegen patches verdwijnen, zonder dat er aanwijsbare verschillen zijn tussen de locaties. De grote variabiliteit in aanplantsucces van jaar tot jaar en van plek tot plek werd bevestigd tijdens de Waddenzee-aanplanten tussen 1991 en 2015 (van Katwijk et al. 2009, van Duren en van Katwijk 2015), maar dit is ook vaak elders in de wereld te zien. Het aanplanten in meerdere jaren is dus om twee redenen van belang: risicospreiding, en het opbouwen van een kritische massa.
Een heel mooi voorbeeld van het belang van grootschaligheid en herhaald uitzaaien is de succesvolle Groot zeegrasaanplant in Virginia Bays in de Verenigde staten, begonnen in 1999. Hier bleek dat iedere uitzaaiing pas na 4-5 jaar leidde tot een lokale uitbreiding (Orth et al. 2012, McGlathery et al. 2012). Men hanteerde daar ook de strategie om meerdere jaren aan te planten. Vanwege het succes en het belang voor de schelpdierindustrie (die het deels of geheel financiert) is men daar nog jaren mee doorgegaan. In 2010 stond er 1700 ha; in 2015 2500 ha (Reynolds et al. 2016). De kritische massa is in dit gebied duidelijk bereikt want de populatie breidt zich nog steeds verder uit, waarbij ook de ecosysteemfuncties zich herstellen, niet alleen de schelpdierpopulaties, maar ook helderder water, fijner sediment, een grote genetische diversiteit aan van zeegras, koolstofopslag en denitrificatie (Reynolds et al. 2016).

Figuur 9.14: Kansenkaart Groot Zeegras
Op basis van een dieptekaart van het Grevelingenmeer is door RWS (Dick de Jong) een indicatieve kansenkaart voor Groot zeegras geproduceerd (9.14). Deze geeft een goede indicatie van de gebieden waar dit meer zeer kansrijk is voor herintroductie van Groot zeegras (tussen -1 en -3 m diepte).
Samenvattend
Het Grevelingenmeer herbergde tijdelijk een enorme oppervlakte aan zeegrasvelden, tot max. 4600 ha in 1978. Daarna nam het zeegras areaal af na gewijzigd sluisbeheer en in 2000 was het zeegras verdwenen in het Grevelingenmeer. Na analyses blijkt het Grevelingenmeer in potentie nog steeds zeer geschikt voor Groot zeegras, en de reden van verdwijnen in het verleden lijkt weggenomen. Met herintroductieprojecten wordt geprobeerd Groot zeegras weer vaste grond aan de voeten te laten krijgen in het Grevelingenmeer.
9.10 Vissen inclusief vismigratie
###Methode
De ontwikkelingen van de visfauna in het Grevelingenmeer zijn deels bepaald aan de hand van bemonsteringen die door Wageningen Marine Research (o.a. in opdracht van RWS) zijn uitgevoerd en deels gebaseerd op de literatuur. Over de jaren heen is de visfauna voornamelijk bemonsterd met een boomkor en in mindere mate met een pelagisch net (e.g. atoom- en wonderkuil) en fuiken. Om een beeld te krijgen van de ontwikkelingen van de visfauna in het Grevelingenmeer voor en na de afsluiting (tot heden) is er voor gekozen om voornamelijk data te gebruiken afkomstig van de boomkorbemonsteringen zodat vergelijkingen kunnen worden gemaakt tussen jaren. Echter, vangsten met een boomkor geven voornamelijk inzicht op soorten die nabij de bodem zwemmen, hoewel ook pelagische soorten in de vangsten voorkomen. De boomkor geeft daarom niet een compleet beeld van de visfauna in het Grevelingenmeer. Daarom zijn zo goed als mogelijk ook gegevens meegenomen die verzameld zijn d.m.v. andere methodes. Door het verschil in vangst methoden (actief vs. passief) kunnen de resultaten van deze bemonsteringen niet een op een vergeleken worden. Een overzicht van de gebruikte databronnen wordt weergegeven in Tabel 5. De gebruikte bemonsteringsmethodes worden hieronder verder besproken binnen verschillende tijdsperioden.
Tabel 5. Overzicht van databronnen gebruikt voor bepalen ontwikkelingen visbestand Grevelingen
| Jaar | Maand | Instituut | Aantal trekken | Publicaties |
|---|---|---|---|---|
| 1960-1980 | Verspreid over jaar | DIHO | 31-341 trekken/jaar 3 m boomkor, 1 cm maaswijdte | Vaas 1978 |
| 1970 | Sept | RIVO (DFS) | 8 trekken. 3 m boomkor, 2 cm maaswijdte | Doornbos et al. 1986 |
| 1976 | DIHO | 3 vistochten met pelagisch net (2-3 m onder opp.) | Vaas 1978 | |
| 1980-1989 | BUWA | Fuiken, duiken, bijwonen vistochten | Meijer en Waardenburg 1990 | |
| 1982 | Aug | Rijkswaterstaat | 48 trekken (>2 m). 3 m boomkor, 6x6 mm maaswijdte | Vos en Twisk 1990 |
| 1988 | Aug | Rijkswaterstaat | 48 trekken (>2 m). 3 m boomkor, 6x6 mm maaswijdte | Vos en Twisk 1990 |
| 1994 | Aug | BUWA, i.o.v. Rijkswaterstaat | 48 trekken (>2 m, 3 m boomkor) & 32 trekken (0-2 m, 2 m boomkor). Maaswijdte 6x6 mm. 80 trekken pelagisch net | Meijer 1995 |
| 1994-2002 | Stichting Anemoon – MOO project | Waarnemingen sportduikers | Gmelig Meyling et al. 2002 | |
| 2008 | Maart | RAVON (FGRA) i.o.v. WMR/Rijkswaterstaat | 21 trekken. 3 m boomkor, 10-15 mm maaswijdte | Van Kessel et al. 2008 |
| 2011 | April | RAVON (FGRA) i.o.v. WMR/Rijkswaterstaat | 28 trekken. 3 m boomkor, 10-15 mm maaswijdte | Van Kessel et al. 2011 |
| 2013 | April | RAVON (FGRA) i.o.v. WMR/Rijkswaterstaat | 28 trekken. 3 m boomkor, 10-15 mm maaswijdte | Van Kessel et al. 2014 |
| 2016 | Najaar | ATKB | 25 trekken. 2.6 m boomkor | Hop 2016 |
| 2017 | Februari | ATKB (FGRA) i.o.v. WMR/Rijkswaterstaat | 28 trekken. 3 m boomkor. | |
| 2017 | Voorjaar en najaar | ATKB i.o.v. Rijkswaterstaat | 53 trekken (>2 m, 3 m boomkoor, 9 mm maaswijdte) & 52 trekken (0-2 m, 2 m boomkor, 14 mm maaswijdte), 65 trekken pelagische netten. | Hop 2017 |
Periode 1960-1978 voor de afsluiting en daarna
Van 1960 tot 1976 is er door het DIHO (nu NIOZ) in het Grevelingenmeer gevist met een boomkor van 3 m mondopening en 1 cm maaswijdte om te onderzoeken wat de gevolgen waren van de Deltawerken op de flora en fauna in de Zeeuwse en Zuid-Hollandse getijdengebieden (Vaas 1978). Tussen 1960 en 1963 hebben 92 trekken plaatsgevonden verdeeld over 7 vistochten. Een vergelijkbaar aantal trekken heeft plaatsgevonden tussen 1966 en 1969 met 102 trekken verdeeld over 10 vistochten (Vaas 1978). Ook na de afsluiting is er jaarlijks bemonsterd met gemiddeld 63 tot 142 trekken per jaar. In 1976 is er gedurende drie vistochten ook gevist met een groot pelagisch net (op diepten van 2-3 m) op plekken waar de totale diepte meer dan 6 m bedraagt (Vaas 1978). Daarnaast zijn er door het RIVO (Rijksinstituut voor Visserijonderzoek) acht trekken uitgevoerd in najaar 1970 met een boomkor van 3 m mondopening als onderdeel van de Nederlandse Demersal Fish Survey (DFS).
Periode 1979-1998 na de opening van de Brouwerssluis
Van 1979 tot 1980 is intensief visonderzoek verricht in het kader van het ZOWEC (ZOut Water ECologie)-project III (Doornbos 1987). Het DIHO is in oktober 1979 gestart met frequente bemonsteringen gebruik makend van een 3 m boomkor (6 x 6 mm maaswijdte in de staart) in gebieden dieper dan 2 m, en met een 1.9 m boomkor in de ondiepere delen. In augustus 1982 is een bestandsopname uitgevoerd d.m.v. een 3 m boomkor (6 x 6 mm maaswijdte in de staart) waarbij 48 random verdeelde locaties in het Grevelingenmeer zijn bemonsterd (De Vos & Twisk 1990). De bestandsopname werd in 1988 herhaald d.m.v. dezelfde methode en de bemonsteringen van dezelfde gebieden (De Vos & Twisk 1990). De boomkor vangt voornamelijk vissen die nabij een zandige bodem leven. Om meer inzicht te krijgen in de volledige samenstelling van de visfauna is tussen 1980-1989 ook een monitoring-onderzoek uitgevoerd via registratie van door beroepsvissers gebruikte fuiken die zowel benthische als pelagische vissen vangen, en door waarnemingen van duikers (Meijer & Waardenburg 1990, Engelsma et al. 1994). In 1994 heeft weer een bestandsopname plaatsgevonden die bestond uit een herhaling van de in 1982 en 1988 uitgevoerde bestandsopname van bodemvissen met boomkorren (Meijer 1995). Daarnaast zijn ook bemonsteringen van pelagische vissen uitgevoerd d.m.v. een boomkuil. Er zijn drie verschillende boomkuilen (3 m, 6 m zonder en met kneppels) gebruikt afhankelijk van de bemonsterde diepte. Beide bemonsteringen zijn uitgevoerd op 80 locaties (Meijer 1995).
Periode 1999-2005 na de jaarrond openstelling van de Brouwerssluis
In de periode 1999-2005 hebben geen kwantitatieve bemonsteringen plaatsgevonden. Wel zijn er in het kader van het MOO (Monitoring Onderwater Oevers) project waarnemingen gedaan door sportduikers tussen 1994 en 2002 (Gmelig Meyling & Bruyne 2003). Daarnaast zijn er door Van der Linden (2006) interviews uitgevoerd met de schipper en eigenaar van sportvis- en sportduikcharterboot de MS Theo betreft de effecten van het jaarrond openstellen van de Brouwerssluis in 1999 op de samenstelling van vispopulaties in het Grevelingenmeer.
Periode 2006-2016 na het opheffen van de 30 dagen regeling van de Brouwerssluis
Eén van de onderdelen van de biologische monitoring in de zoete Rijkswateren bestaat uit het programma Actieve Monitoring Zoete Rijkswateren waarbij bestandsopnames worden gemaakt d.m.v. kor- en electrovisserij op vaste trajecten in de grote Rijkswateren waaronder het Grevelingenmeer sinds het monitoringsjaar 2007-2008. De bemonsteringen zijn uitgevoerd door Natuurbalans – Limes Divergens en Stichting RAVON in 2008, 2011, 2013 (Van Kessel et al. 2008, 2011, 2014) en door ATKB in 2017. Tussen februari en maart zijn in 2008, 2011, 2013 en 2017 respectievelijk 23, 28, 28 en 28 trekken uitgevoerd met een 3 m brede boomkor (kleinste maaswijdte = 10-15 mm gestrekt) over een afstand van circa 1000 m.
Daarnaast heeft ATKB in 2017 zowel in het voor- als najaar een extra bemonstering uitgevoerd waarbij in ondiepe delen (0-2 m) gevist is met een 2 m boomkor (kleinste maaswijdte = 9 mm in zak) over een lengte van 500 m (52 trekken), en in diepere delen met een 3 m boomkor met een maaswijdte van 14 mm in de zak (53 trekken) (Hop 2017).
Bij het bemonsteren van vissoorten in de waterkolom is een atoomkuil (5 x 5 m, maaswijdte 10 mm in de zak) toegepast in diepe delen (>5 m) en een kleine wonderkuil (3 x 0.8 m, maaswijdte 12 mm in de zak) in de ondiepe delen (<5 m), beide over een afstand van 1000 m. Voor zowel de boomkor, atoomkuil en wonderkuil worden vangstrendementen toegepast (Hop 2017). Daarnaast is er door ATKB in november 2016 een visstand onderzoek uitgevoerd in opdracht van recreatieschap Zuidwestelijke Delta om zowel de bodem gebonden- als pelagische vis goed in beeld te brengen (Hop 2016).
Verspreid over het Grevelingenmeer zijn 25 trekken (500 m) met een 2.6 m boomkor genomen en 10 trekken (1000 m) met een atoomkuil welke zich bevond in de bovenste, dan wel onderste laag (Hop 2016). De maaswijdte van deze twee tuigen is onbekend.
Overige analyses
Naast een jaarlijks gemiddelde is ook gekeken hoe soorten, aantallen en biomassa zich verdelen over het Grevelingenmeer, gebruik makend van de data uit 1970, 2008, 2011, 2013 en 2017, verzameld door RIVO/Wageningen Marine Research (Tabel 5). Om deze horizontale gradiënt in kaart te brengen is het Grevelingenmeer opgedeeld in acht gebieden op basis van de vakindeling van Wetsteijn (2011), welke is gebruikt voor onderzoek naar stratificatie en zuurstof deficiëntie. Deze vakindeling is later ook gebruikt door Hop (2016, 2017) om de verspreiding van diverse vissoorten weer te geven (Figuur 9.15).

Figuur 9.15: Vakindeling Grevelingenmeer (Wetsteijn 2011, Hop 2017)
De vissoorten zijn verder onderverdeeld in een aantal groepen die corresponderen met relevante kenmerken van een deel van het specifieke watersysteem (e.g. chloridegehalte, isolatie/verbinding). Deze groepen, ecologische gilden, kunnen een indicatie geven over de verandering in een systeem. Vissoorten zijn ingedeeld in ecologische gildes volgens de indeling van Elliott & Hemingway (2002) voor estuaria, deze indeling wordt ook gebruikt voor de KRW:
- Diadrome soorten (CA): soorten die migreren tussen zee en rivier en het estuarium als trekroute gebruiken en soms ook (tijdelijk) als opgroeigebied;
- Estuarien residente soorten (ER): soorten die hun totale levenscyclus in het estuarium kunnen doorlopen;
- Mariene juvenielen (MJ): mariene soorten waarvan de jonge exemplaren kunnen opgroeien in een estuarium;
- Mariene seizoensgasten (MS): mariene soorten die in een vast seizoen een estuarium kunnen bezoeken;
- Mariene gast (MA): zeesoort zonder speciale behoefte aan een estuarium, bezoekt onregelmatig;
- Zoetwatersoorten (FW): soorten die hun levenscyclus voornamelijk in zoetwater kunnen doorlopen maar, afhankelijk van de zouttolerantie van elke soort, soms ook in brakwater gevonden worden.
Daarnaast zijn de vissoorten ook onderverdeeld in groepen volgens hun voorkeur voor voedselinname, ofwel voedselgildes. Elke voedselgilde vertegenwoordigt dus een deel van de vis gemeenschap met een bepaalde voedsel keuze. Deze voedselgildes kunnen gebruikt worden om de structuur en het functioneren van estuariene vispopulaties te bepalen. Vissoorten zijn ingedeeld in de volgende groepen: Benthivoren, bentho-piscivoren, piscivoren en planktivoren.
9.10.1 Resultaten ontwikkeling van vis
Aan de hand van literatuuronderzoek en data verzameld door RIVO/Wageningen Marine Research wordt een overzicht gegeven van de ontwikkelingen van biomassa (g/m2) en het bemonsterde aantal soorten in de jaren dat een bestandsopname heeft plaatsgevonden (Figuur 9.16). Wel moet opgemerkt worden dat het aantal trekken, trekduur, bemonsteringslocaties en bemonsteringsmethodes verschillen tussen jaren (Tabel 5) waardoor biomassa en het aantal gevangen soorten niet altijd direct tussen jaren vergeleken kan worden. Wel kan gezegd worden dat biomassa hoger was voor de afsluiting van het Grevelingenmeer dan erna en lijkt de soortenrijkdom ook wat te zijn afgenomen na de sluiting. Dit wordt hieronder verder besproken.

Figuur 9.16: Totale biomassa (gram per m2, linker y-as) en aantal soorten gevangen per jaar (rechter y-as) gebaseerd op boomkorbemonsteringen in het Grevelingenmeer tussen 1960 en 2017.
Periode 1960-1970 voor de afsluiting (boomkorbemonstering) Tussen 1960 en 1963 bestond de bodemvisfauna voornamelijk uit mariene soorten; zeesoorten die het gebied onregelmatig bezoeken, bijvoorbeeld als opgroeigebied voor juveniele soorten of als foerageergebied voor volwassen vissen, maar geen speciale behoefte hebben aan een estuarium (Hartgers et al. 2001). Zo gebruikte o.a. de jonge schol (Pleuronectes platessa) en tong (Solea solea) de Grevelingen als opgroei habitat en trokken soorten zoals spiering (Osmerus eperlanus) en fint (Alosa fallax) in het voorjaar stroomopwaarts om in het zoete rivierwater te paaien (Vaas 1978). Een andere migrerende soort die gebruik maakte van het gebied was de paling (Anguilla anguilla). Jonge paling (glasaal) trok in het voorjaar het zoete water op om daar op te groeien. Na 5 tot 20 jaar keert de paling in het najaar terug naar zee om zich in de Sargassozee voort te planten (schieraal) (Hartgers et a. 2001 en referenties daarin). Andere soorten die gebruik maakten van de Grevelingen als estuarium zijn de driedoornige stekelbaars (Gasterosteus aculeatus), brakwatergrondel (Pomatoschistus microps), puitaal (Zoarces viviparus) en de zeedonderpad (Myxocephalus scorpius*) welke zich zowel in kustwateren als estuaria met hoge zoutgehaltes kunnen voortplanten (Vaas 1978). Gedurende deze periode vertegenwoordigde de tong het grootste aandeel (56.5 kg) in de totale biomassa (120 kg) gevolgd door schol (26.5 kg) en de zeedonderpad (7 kg). Het aantal soorten tussen 1960 en 1963 bedroeg 27 (Vaas 1978).
De aanleg van de Grevelingendam (1958-1965) zorgde ervoor dat de Grevelingen afgesloten werd van de aanvoer van rivierwater en vormde een barrière voor mariene soorten zoals paling, spiering en fint om verder stroomopwaarts te trekken naar paai- en opgroeigebieden. Desondanks bleef de soortensamenstelling van de bodemvisfauna vrijwel gelijk met 31 soorten in de periode na de bouw van de Grevelingendam (1966-1969). Wel bleek dat tong in deze periode niet meer de voornaamste bodemvis was en vervangen werd door schol en schar (Limanda limanda).

Figuur 9.17: Biomassa (kg per ha) en aantallen (n per ha) vissoorten in verschillende vakken in de Grevelingen in het najaar 1970, gebaseerd op N=8 trekken. In vakken 1, 3 en 7 zijn geen trekken genomen. De errorbars geven de standaard deviatie weer omdat in sommige vakken meerdere trekken zijn genomen.
De ruimtelijke verdeling van biomassa en aantallen vis in het najaar van 1970 is weergegeven in Figuur 9.17. De genomen trekken (N=8) vielen binnen vijf vakken. De grootste dichtheden komen voor in vak 8 en worden voornamelijk veroorzaakt door het aantal gevangen schollen (N=169/ha), scharren (N=153-157/ha) en tongen (N=127/ha) per trek. De trekken binnen vak 8 zijn genomen ten noordoosten en noordwesten van het eiland Hompelvoet, het westelijk deel van de Grevelingen. De hoge dichtheid in vak 4 wordt voornamelijk veroorzaakt door grondels (N=133/ha, Pomatoschistus sp.), scharren (N=109/ha) en tongen (N=128/ha). Schollen (9.9 kg/h) zorgen voor een hogere biomassa in vak 4. Het aantal verschillende soorten dat nabij de bodem is aangetroffen is het hoogste in vak 8 (N=9) en het laagst in vakken 2 en 4 (N=6).
Ecologische gildes
Om inzicht te krijgen in hoe groepen van vissen veranderen met specifieke kenmerken van het watersysteem (bv. connectiviteit) is ook gekeken naar ecologische gildes. Dit is alleen gedaan voor het jaar 1970 waarvoor data beschikbaar waren (Figuur 9.19). Het grootste aantal soorten behoort tot het gilde van de mariene juvenielen, welke ook de biomassa domineert. Dit zijn de soorten schar, schol, tong, koornaarvissen (Atherina sp.), haring (Clupea harengus), zeebaars (Dicentrarchus labrax), smelt (Hyperoplus marinus), wijting (Merlangius merlangus) en steenbolk (Trisopterus luscus). De verspreiding is relatief gelijk over de bemonsterde vakken. De estuarien residente soorten spelen ook een belangrijke rol in de dichtheid maar niet zo zeer in de biomassa. Aangetroffen soorten zijn bot (Platichthys flesus), harnasmannetje (Agonus cataphractus), zeenaalden (Syngnathus sp.) en grondels. Alleen in vak 2 zijn diadrome soorten aangetroffen zoals paling en spiering. Mogelijk omdat de constructie van de Brouwersdam (1962-1971) al bijna klaar was en er al een fysieke barrière aanwezig was waardoor diadrome soorten niet naar zee konden trekken.

Figuur 9.18: Aandeel van biomassa (%) voor elke ecologische gilde (CA=diadrome soorten, ER=estuarien residente soorten, MA=Mariene gast, MJ=Mariene juvenielen, MS=Mariene seizoensgasten) in verschillende vakken in de Grevelingen in het najaar 1970.

Figuur 9.19: Aandeel van dichtheid (%) voor elke ecologische gilde (CA=diadrome soorten, ER=estuarien residente soorten, MA=Mariene gast, MJ=Mariene juvenielen, MS=Mariene seizoensgasten) in verschillende vakken in de Grevelingen in het najaar 1970.

Figuur 9.20: Aandeel van biomassa (%) voor elk voedselgilde in verschillende vakken in de Grevelingen in het najaar 1970.

Figuur 9.21: Aandeel van dichtheid (%) voor elk voedselgilde in verschillende vakken in de Grevelingen in het najaar 1970.
Voedselgildes
Ook is voor het jaar 1970 gekeken naar een indeling in voedselgildes (Figuur 9.21). Het grootste aantal soorten behoort tot de benthivoren met soorten zoals schol, schar, tong, bot, slakdorf (Liparis liparis), steenbolk, botervis (Pholis gunnellus) en harnasmannetje. De soorten slakdorf, spiering, en puitaal werden alleen in het westelijke deel van de Grevelingen aangetroffen. Wat biomassa betreft, waren ook soorten die behoren tot de planktivoren belangrijk met haring en grondel in het westelijk deel (vak 2 & 8) en zeenaalden in het oostelijk deel (vak 4-6). De soorten behorende tot de bentho-piscivoren, zoals paling en zeedonderpad, leverden maar een kleine bijdrage aan de biomassa in vakken 4 tot 6.
Periode 1971-1978 na de afsluiting (boomkorbemonstering)
Het afsluiten van de Grevelingen door de aanleg van de Brouwersdam in de zomer van 1971 had zowel directe als indirecte gevolgen voor vis. Nabij de Brouwersdam stierven grote hoeveelheden harders en gepen in december 1971 en januari 1972 toen het water te koud werd (Waardenburg 1998). Kennelijk konden deze soorten geen manier vinden om te ontsnappen naar de Noordzee. Na de afsluiting daalde het aantal soorten sterk van 32 tussen 1960 en 1969 (Vaas 1978) naar 26 in 1971. Aangezien het aantal soorten sterk werd beïnvloed door de trefkans van zeldzame en weinig frequente soorten heeft Vaas (1978) ook de aantallen vergeleken over langere perioden van 4 jaar. Op deze manier is te zien dat in de eerste jaren na de afsluiting het aantal soorten relatief gelijk bleef en dat het aantal soorten pas in latere jaren afnam (vanaf ~1974).
Vaas (1978) heeft de soortensamenstelling nader geanalyseerd en verdeelde de soorten in de volgende groepen:
- Soorten die onmiddellijk na de afsluiting verdwenen en later ook niet meer gevangen werden zoals de dwergtong (Buglossidium luteum), pitvissen (Callionymus sp.), zandspiering (Ammodytes tobianus), rode poon (Chelidonichthys lucerna), adderzeenaald (Entelurus aequoreus), geep (Belone belone) en fint. Dit zijn voornamelijk soorten die al met een lage frequentie en in geringe aantallen gevangen werden.
- Soorten die tijdens de periode van de afsluiting verdwenen zoals tong, tongschar (Microstomus kitt), spiering, harnasmannetje, zeenaald, kabeljauw (Gadus morhua), vijfdradige meun (Ciliata mustela) en de horsmakreel (Trachurus trachurus). Het gaat hier om vissen die vanuit zee het Grevelingenmeer inzwemmen maar zich niet in de afgesloten Grevelingen kunnen voortplanten. Deze soorten hebben zich iets langer weten te handhaven en speelden voor de afsluiting een belangrijke rol in de totale biomassa (voornamelijk tong en spiering).
- Soorten die voor de afsluiting ontbraken, na de afsluiting gevangen werden, maar in 1976 ontbraken. Het zijn soorten die incidenteel in de vangsten voorkomen na de afsluiting zoals de glasgrondel (Aphia minuta), Europese zeebaars en vorskwab (Raniceps raninus).
- Soorten die voor de afsluiting ontbraken en in de periode daarna gevangen werden (sowieso tot 1976) zoals tarbot (Scophthalmus maximus), griet (Scophthalmus rhombus), ansjovis (Engraulis encrasicolus) en de zwarte grondel (Gobius niger). Het is waarschijnlijk dat de eerste drie soorten niet geheel afwezig waren voor de sluiting maar nadat ze eenmaal binnen waren gedrongen in het afgesloten meer ze zich op hun gemak voelden. De zwarte grondel is wel een nieuwe soort in het Grevelingenmeer. Deze soort werd ook aangetroffen in het Veerse Meer (Vaas 1970), drie jaar na het ontstaan, en heeft zich in 1966 ontwikkeld tot een grote populatie (Vaas 1978). Hetzelfde gebeurt in het Grevelingenmeer.
Als de 12 meest dominante soorten voor de afsluiting worden vergeleken met de 12 meest dominante na de afsluiting zijn 8 soorten hetzelfde gebleven: schol, schar, bot, dikkopje (Pomatoschistus minutus), paling, sprot (Sprattus sprattus), haring en puitaal, alhoewel binnen deze groep sterke verschuivingen hebben plaatsgevonden (Vaas 1978). Tong, spiering, zeedonderpad en wijting zijn na de afsluiting vervangen door de brakwatergrondel, zwarte grondel, koornaarvissen en driedoornige stekelbaars. Sprot nam ook sterk toe. Waarschijnlijk omdat een grote populatie sprot zich in het Grevelingenmeer bevond wanneer deze werd afgesloten en zich daarna in het meer heeft voortgeplant. Echter, de sprot en koornaarvissen namen ook weer vrij snel af, mogelijk door de verzoeting van het Grevelingenmeer wat gevolgen had voor de reproductie (Doornbos 1982). Het vermogen tot voortplanting is bepalend geweest voor de ontwikkeling van de visfauna in het Grevelingenmeer. De soorten die zich niet in het afgesloten meer kunnen voortplanten, zijn vrijwel verdwenen. De grondel en stekelbaars planten zich wel in het meer voort. Het aantal soorten in het Grevelingenmeer bleef echter dalen tot 1975 en steeg in 1976 met één soort (zwarte grondel). De afsluiting van de Grevelingen zorgde zowel voor een daling in diversiteit als voor een verandering in de samenstelling van de visfauna van voornamelijk grote naar kleinere soorten (Doornbos 1982).
De biomassa nam voor de meeste soorten ook sterk af na de afsluiting behalve voor schol welke gelijk bleef. In de jaren 1972 en 1973 steeg de biomassa van de schol sterk doordat de jonge dieren sterk groeiden en zich van de ondiepe naar de diepere delen verplaatsten. In 1974 ging de biomassa van schol weer achteruit omdat de scholpopulatie zich niet voorplant in de afgesloten Grevelingen. De totale biomassa bleef echter gelijk omdat andere vissoorten zich hersteld hadden zoals bot en paling, en in mindere mate grondel en puitaal. Dit had tot gevolg dat de totale biomassa alsnog steeg (Vaas 1978).
Periode 1979-1998 na de opening van de Brouwerssluis (boomkorbemonstering, fuikenbemonstering en monitoring door duikers).
Met het jaarrond openzetten van de spuisluis steeg het zoutgehalte en nam het aantal waargenomen soorten toe van 20 in 1977 en 1978 tot 26 in 1979 (Figuur 9.22) (gebaseerd op boomkor bemonsteringen). Echter in 1980 daalde het aantal gevangen soorten weer naar 22 toen alleen water vanuit de Noordzee werd ingelaten in de winter periode. De ‘nieuwe’ soorten in 1979 zijn voornamelijk zomergasten zoals geep, horsmakreel en diklipharder (Chelon labrosus), en werden in 1980 niet meer in de boomkorvangsten aangetroffen (Doornbos 1982). Proeffuiken vingen een week na het inlaten van Noordzeewater ook nog andere soorten zoals de fint, makreel (Scomber scombrus), zeebaars, zeeforel (Salmo trutta) en zeeprik (Petromyzon marinus) (Philippart & Meijer 1982). Daarnaast gaven waarnemingen van rijpe sprot vrouwtjes en larven aan dat de soort zich weer voortplanten in het meer (Doornbos 1982). Soorten zoals de zwarte grondel, brakwatergrondel en het dikkopje namen toe tussen 1978 en 1980 (Doornbos 1982). Ook de scholpopulatie kreeg een boost. De schollen die nog aanwezig waren in het meer (beperkt tot een aantal volwassen exemplaren) konden het meer verlaten in de winter om naar hun paaiplaatsen te migreren in de Noordzee en larven konden het meer binnenkomen in het voorjaar (Doornbos 1982). Het succes van de intrek van de schollarven was echter wel afhankelijk van het sluisbeheer. Vanaf 1980 was de spuisluis alleen open van half oktober tot maart. Dit beheer had tot gevolg dat volwassen schollen in de winter naar de Noordzee konden trekken (Steinmetz & Slothouwer 1979, Doornbos 1981) maar dat de pelagische larven vaak niet het Grevelingenmeer in konden komen omdat deze pas in maart-april bij het estuarium aankwamen (Rijnsdorp et al. 1985, Van der Veer 1985), dan was de sluis al gesloten. Dit gold tevens ook voor botlarven welke pas in april aankwamen (Van der Veer 1985). In 1985 bleef de sluis echter wel lang genoeg open, tot 22 maart, waardoor een grote hoeveelheid schollarven zich heeft kunnen vestigen in het meer en (voorlopig) heeft kunnen voorkomen dat de scholpopulatie verder afnam (Doornbos 1987). Uit de bovenstaande gegevens blijkt dat het spuibeheer een sterke invloed heeft gehad op het aantal soorten dat voorkwam in het Grevelingenmeer.

Figuur 9.22: Aantal vissoorten gevangen met de 3 m boomkor in de diepere delen (>5 m) van het Grevelingenmeer en de Shannon-Weaver diversiteitsindex (uit Doornbos 1987).
In de periode 1980-1989 is zowel fuikenmonitoring (jaarlijks) als boomkormonitoring (alleen in de jaren 1982, 1988 en 1994) uitgevoerd om het visbestand in kaart te brengen. Met fuiken worden meer vissoorten aangetroffen met een wat andere soortensamenstelling dan wanneer er bemonsterd wordt met een boomkor. Tabel 6 geeft aan welke soorten zijn gevangen en in welke frequentie. Tijdens de fuikenmonitoring zijn over de hele periode totaal 58 vissoorten aangetroffen (met 44-51 soorten per jaar). Opgemerkt dient te worden dat harders als één soort zijn geregistreerd en dat er geen onderscheid gemaakt is tussen zalm (Salmo salar) en zeeforel (Meijer & Waardenburg 1990).
Tabel 6. Gevangen vissoorten d.m.v. fuiken in het Grevelingenmeer tussen 1980-1989, ingedeeld in presentie-categorieën (Meijer & Waardenburg 1990). De vrij zeldzame soorten kunnen mogelijk ondervertegenwoordigd zijn door een grotere ontsnappingskans uit een fuik.
| Presentie-categorieën | Omschrijving | Vissoorten |
|---|---|---|
| Zeer algemeen | In 50-100% van alle vangsten aanwezig | Paling, bot, schol, puitaal, haring, zeedonderpad, griet |
| Algemeen | In 25-50% van alle vangsten aanwezig | Schar, geep, tong, tarbot, koornaarvissen, kabeljauw, wijting |
| Vrij algemeen | In 10-25% van alle vangsten aanwezig | Zwarte grondel, brakwatergrondel, dikkopje, vijfdradige meun, snotolf |
| Vrij zeldzaam | In 1-10% van alle vangsten aanwezig | Botervis, zeenaald, harder, zeebaars, fint, steenbolk, pitvis |
| Zeldzaam | In <1% van alle vangsten aanwezig | Zeeprik, rivierprik, pollak, sardien |
De meest voorkomende soorten in de fuikvangsten waren paling, schol en bot. Kleine paling (30-40 cm) wordt voornamelijk met de boomkor gevangen en individuen groter dan 30 cm hoofdzakelijk met de pelagische trawl. Dit wijst op een pelagische levenswijze gedurende de zomer waar paling onder andere jaagt op scholen sprot (Doornbos 1985). De paling is verspreid over het hele meer aanwezig (Meijer & Waardenburg 1990). Paling wordt ook beroepsmatig gevist en vissers zagen vanaf ongeveer 1988 een afname van het aantal gevangen palingen, ervan uitgaand dat de visserij-inspanning gelijk gebleven is (Engelsma et al. 1994). Deze trend lijkt zich door te hebben gezet in het begin van de jaren negentig. Mogelijke oorzaken van de teruggang in de paling populatie zijn het hoge zoutgehalte in het Grevelingenmeer welke de intrek van glasaal beïnvloed en het verdwijnen van zeegrasvelden en de mogelijke afname van Japans bessenwier (Engelsma et al. 1994). Het verdwijnen van zeegras en de afname van bessenwier valt samen met de achteruitgang van de paling (Engelsma et al. 1994). Een directe link is nog niet gelegd. Andere soorten zoals de zeedonderpad, botervis, zwarte grondel, vorskwab en puitaal zijn kenmerkende bewoners van hard substraat zoals dat wordt aangetroffen langs dijken bijvoorbeeld. Het grootste aantal soorten wordt waargenomen rondom de Brouwerssluis en is daar hoger dan in andere delen van het Gevelingenmeer (Engelsma et al 1994). Dit is in overeenkomst met het macrozoöbenthos waar de diversiteit aan soorten en de totale biomassa ook hoger zijn in het westelijke deel (Mulder et al. 2019).
Het aantal vissoorten gevangen met de boomkor is in alle jaren vergelijkbaar met 19 soorten in 1982, 18 soorten in 1988 en 19 soorten in 1994. De belangrijkste soorten waren vooral de zwarte grondel, brakwatergrondel, dikkopje, jonge schol en de pelagische soorten sprot, haring en koornaarvissen (Meijer 1995).
De brakwatergrondel komt voornamelijk voor in de litorale zone (0-2 m) en het dikkopje en zwarte grondel in de sublitorale zone (>2 m). De totale biomassa van grondels in het sublitoraal is gelijk gebleven tussen 1982 en 1988, alhoewel in 1982 het dikkopje in biomassa domineerde en in 1988 juist de zwarte grondel. In 1994 wordt weer een omkering van rangorde gevonden en domineert het dikkopje weer. Bedacht moet worden dat het een moment opname betreft en dat de biomassa van grondels gedurende het jaar sterk fluctueert.
Zoals eerder genoemd, is de populatie van platvissen sterk afhankelijk van de intrek van larven in het voorjaar aangezien ze zich niet voorplanten in het Grevelingenmeer. De intrek wordt o.a. bepaald door het sluisbeheer en de sterkte van de Noordzee-populatie. In 1979, 1985 en 1994 vond een grote intrek van schollarven plaats, o.a. doordat de sluis in 1994 tot begin mei heeft open gestaan. Desondanks zijn de aantallen en biomassa van schol in 1994 sterk afgenomen ten opzichte van 1988. Het aantal oudere schollen is sterk afgenomen en er wordt gesuggereerd dat deze mogelijk zijn weggetrokken in het najaar naar de Noordzee (Meijer 1995). Bot komt in veel kleinere aantallen voor dan schol maar de populatie neemt wel toe van 1982 tot 1994. Mogelijk door een betere intrek van larven in 1994 of door een dalende scholpopulatie. Net als bij de scholpopulatie daalt de biomassa van bot sneller dan de aantallen en is het aandeel van de oudere exemplaren dus hier ook gedaald (Meijer 1995).
Wat betreft de pelagische soorten (alleen in 1994 bemonsterd) lijkt er een scheiding te zijn waarbij koornaarvissen voornamelijk voorkomen in de ondiepe zones (<5 m) en de haring en sprot juist in de diepere delen van het meer. Deze drie soorten vertegenwoordigden in 1994 een totale biomassa van ruim 100 ton waarvan sprot het grootste deel vertegenwoordigde (79%) gevolgd door haring (12%) en koornaarvissen (~9%). Het merendeel bestond uit 0-jarige individuen. Desondanks vertegenwoordigde de pelagische vissen het grootste deel van de biomassa (68%) in vergelijking met grondels of platvissen (Meijer 1995). De driedoornige stekelbaars ontbreekt vrijwel in 1994 met slechts enkele exemplaren in de diepere delen van het meer. Deze soort was vroeger in de zeegrasvelden van belang (Doornbos 1985). Het Grevelingenmeer bevatte namelijk grote zeegrasvelden welke in de ondiepere delen (<2 m) voorkwamen en een goede leefomgeving bood voor kleine vissoorten zoals de brakwatergrondel, dikkopje, en paling, maar ook fungeerde als paaigebied voor stekelbaarzen. Echter na 1978, wanneer de grootste bedekking werd bereikt (4400 ha), nam het zeegrasareaal sterk af tot 1427 ha en speelde rond 1994 geen rol meer. Vissoorten die gebonden zijn aan deze vegetatie spelen nu geen grote rol meer in vergelijking met vroeger (De Vos & Twisk 1990, Meijer 1995).
Een andere verandering die plaatsvond in het Grevelingenmeer was de sterke toename van het muiltje (Crepidula fornicata). Deze toename van hard substraat is voordelig voor grondels welke hun eieren erop af kunnen zetten. Er wordt dan ook gesuggereerd dat een verband tussen de toename van schelpdieren waaronder het muiltje en grondels aannemelijk lijkt (Meijer 1995). De verandering van een zacht- naar een hardsubstraat bodem kan grote gevolgen hebben voor soorten die op of in het zacht-substraat leven zoals platvissen. Meijer (1995) suggereert dat dit mogelijk de afname van de platvissen voor een deel kan verklaren.
Monitoring door duikers suggereert dat het openstellen van de Brouwerssluis positieve effecten heeft gehad op de diversiteit aangezien duikers een toegenomen soortenrijkdom van vissen hebben waargenomen (Gmelig Meyling & Bruyne 2003). Noordzee-vissoorten zoals makreel, geep, wijting, steenbolk, jonge kabeljauw, rode poon en rog worden steeds vaker en in grotere hoeveelheden gevangen maar ook soorten zoals pitvis, tong, zeedonderpad en zeebaars zijn toegenomen (Van de Linden 2006, Gmelig Meyling & Bruyne 2003). Daarnaast worden er zowel grote als kleine schollen gevangen. Het jaarrond openstellen heeft er ook voor gezorgd dat schollarven nu altijd het Grevelingenmeer in kunnen trekken, in tegenstelling tot het spuibeheer van voor 1999 (Van der Linden 2006). Wel wordt er aangegeven dat tijdens de sluiting van 30 dagen in het najaar, er niks tot weinig wordt gevangen in het meer. Pas een week na de openstelling gaan de vangsten weer omhoog (Van de Linden 2006).
*Periode 2006-2017 na het jaarrond openen van de Brouwerssluis In 2008, 2011, 2013 en 2017 zijn boomkorbemonsteringen uitgevoerd. Het mogelijke effect op het visbestand van het opheffen van de 30 dagen regeling in 2005 is moeilijk te bepalen omdat er weinig tot geen bemonsteringen hebben plaatsgevonden rondom deze periode. Echter het lijkt aannemelijk dat het jaarrond openzetten van de spuisluis voordelige gevolgen gehad zal hebben voor soorten die in het najaar het Grevelingenmeer willen verlaten of binnentrekken, wat eerder bemoeilijkt werd door de 30 dagen sluiting tussen september en december.
###Biomassa en dichtheid
Tijdens de bemonstering in 2008 zijn 23 verschillende soorten aangetroffen. De soorten die in de grootste aantallen zijn aangetroffen zijn het dikkopje (100-1300 n/ha) en de zwarte grondel (100-700/ha), gevolgd in lagere dichtheden door schol, tong en schar. De biomassa wordt gedomineerd door schol, tarbot, griet en tong (10-30 kg/ha). De verdeling van biomassa en aantallen vissoorten over het Grevelingenmeer in voorjaar 2008 zijn weergegeven in Figuur 28. De hoogste biomassa en dichtheden komen voor in vak 6 en worden voornamelijk veroorzaakt door het dikkopje (0.1 kg/ha, 100 n/ha) en de zwarte grondel (0.1-2.3 kg/ha, 300-700 n/ha). In vak 1 en 2 dragen ook andere soorten bij aan een verhoogde biomassa zoals griet, tarbot, schol en tong (10-30 kg/ha). In 2011 zijn tijdens de bemonstering 13 soorten aangetroffen, een aanzienlijk lager aantal dan in 2008. De soorten die in de grootste aantallen zijn aangetroffen, zijn de zwarte grondel, schol, brakwatergrondel, puitaal en het dikkopje. Qua biomassa domineren de zeedonderpad en de schol. De verdeling van biomassa en aantallen vissoorten over het Grevelingenmeer in voorjaar 2011 zijn weergegeven in Figuur 9.23. De dichtheid is relatief hetzelfde per vak alhoewel in vak 2, 4 en 7 er zich enigszins grotere aantallen vis bevinden. In vak 2 zorgen voornamelijk schol (90 n/ha) en zwarte grondel (25 n/ha) voor hogere dichtheden, in vak 4 puitaal (40 n/ha), brakwatergrondel (32 n/ha) en grote zeenaald (25 n/ha) en in vak 7 zorgt voornamelijk de zwarte grondel (36 n/ha) voor een hogere dichtheid gevolgd in lagere dichtheden door dikkopje, puitaal, brakwatergrondel en botervis (~6 kg/ha). In vak 2 draagt voornamelijk schol (2-3 kg/ha) bij aan een hogere biomassa.

Figuur 9.23: Biomassa (kg/ha, links) en aantallen (n/ha, rechts) vissoorten in verschillende vakken in het Grevelingenmeer in het voorjaar 2008, 2011, 2013 en 2017. De errorbars geven de standaard deviatie weer omdat in sommige vakken meerdere trekken zijn genomen.
Tijdens de bemonstering in 2013 zijn 13 verschillende vissoorten aangetroffen, een dezelfde aantal soorten als in 2011 maar met een wat andere samenstelling. Soorten die wel in 2011 en niet in 2013 zijn aangetroffen zijn de zeedonderpad, brakwatergrondel, zandspiering en ansjovis. Tevens is in 2011 de kleine zeenaald aangetroffen en in 2013 alleen de grote zeenaald. De meeste van deze soorten (m.u.v. de brakwatergrondel) zijn in kleine aantallen gevangen en de trefkans om deze soorten bij iedere bemonstering te vangen is waarschijnlijk laag. De soorten die in 2013 in hoge aantallen voorkwamen, zijn het dikkopje, zwarte grondel, schol en botervis, gevolgd door puitaal en sprot in lagere aantallen. De soorten die de grootste bijdrage leveren aan de totale biomassa zijn de zwarte grondel, schol, dikkopje en botervis. De verdeling van biomassa en aantallen vissoorten in voorjaar 2013 zijn weergegeven in Figuur 9.23. De dichtheid is relatief gelijk verspreid over het meer met in vak 2 en 4 uitschieters welke worden veroorzaakt door het dikkopje (200-600 n/ha). Daarnaast is de biomassa het hoogste in het zuidwestelijk deel van het Grevelingenmeer (vak 1-4), wat voornamelijk veroorzaakt wordt door het dikkopje (tot 1.0 kg/ha) en de schol (tot 0.67 kg/ha) en in kleinere hoeveelheden door botervis en puitaal (0.05-0.08 kg/ha).
In het voorjaar van 2017 (toen de Flakkeese Spuisluis net geopend was) zijn, d.m.v. boomkorbemonsteringen, 17 soorten aangetroffen. Soorten die in 2017 zijn aangetroffen maar niet in 2011 en 2013 zijn zeebaars, koornaarvissen, adderzeenaald en haring. De zeebaars en koornaarvissen waren voor het laatst aangetroffen in 2008. De soorten die in 2017 in hoge aantallen voorkwamen, zijn haring (1245-2900 n/ha), dikkopje (1100-10,000 n/ha) en sprot (1300 n/ha). Deze hoge aantallen kwamen voornamelijk voor in vak 1 en 2 (Figuur 28). De hoge biomassa in vak 1 werd voornamelijk veroorzaakt door sprot (11 kg/ha), haring (25 kg/ha) en het dikkopje (31 kg/ha).
9.10.2 Ecologische gildes
Het grootste aantal soorten behoort tot de gilde estuarien residente soorten, welke in alle jaren zowel in biomassa als in aantallen domineren. Dit zijn over het algemeen (met enkele uitzonderingen) soorten die hun hele levenscyclus in het Grevelingenmeer kunnen voltooien zoals de zwarte grondel, dikkopje, brakwatergrondel, puitaal en in mindere mate de zeenaald. Er heeft een duidelijke verschuiving plaatsgevonden in verhoudingen in vergelijking met 1970 (Figuur 9.18). In 1970 werd de biomassa bijna compleet gedomineerd door mariene juvenielen terwijl deze nu nog maar een kleine rol spelen en dan voornamelijk in het westelijk deel van het Grevelingenmeer (Figuur 9.24). De mariene juvenielen bestaan in 2008, 2011 en 2013 vooral uit tong, schol en schar, en in 2017 voornamelijk uit haring, zeebaars en koornaarvissen. In 2017 is ook de adderzeenaald (mariene gast) aangetroffen in vak 4 welke een duidelijke bijdrage levert aan de totale biomassa. Er worden heel beperkt diadrome soorten aangetroffen in de bemonsteringen. Zo werd in 2008 één paling gevangen en werden in 2013 twee spieringen en één driedoornige stekelbaars gevangen. Echter in 2017 zijn een groot aantal driedoornige stekelbaarzen gevangen, voornamelijk in het oostelijk deel van het Grevelingenmeer (Figuur 9.24). Het kan zijn dat een deel van de populatie in het voorjaar via de Brouwerssluis het Grevelingenmeer optrekt (Hop 2017), net als vroeger (Doornbos 1985). Er moet echter opgemerkt worden dat dit ook een resultaat kan zijn van de verhoogde bemonsteringsinspanning in 2017.
Hieronder wordt de verhouding van de verschillende gildes weergegeven per vak voor biomassa en dichtheid in voorjaar 2008, 2011, 2013 en 2017 (Figuur 9.24).

Figuur 9.24: Aandeel van biomassa (procent, links) en dichtheid (procent, rechts) voor elke ecologische gilde (CA=diadrome soorten, ER=Estuarien residente soorten, MA=Mariene gast, MJ=Mariene juvenielen, MS=Mariene seizoensgasten) in verschillende vakken in het Grevelingenmeer in voorjaar 2008, 2011, 2013 en 2017.
###Voedselgildes
Waar de benthivoren in aantallen en biomassa domineerden in 2008, domineren de planktivoren in 2017 (Figuur 9.25). De benthivoren in 2008 bestonden voornamelijk uit de zwarte grondel, tong, schol en schar, en de planktivoren uit het dikkopje en sprot. In 2011 en 2013 werd de tong nauwelijks gevangen en werden benthische soorten zoals puitaal en botervis meer aangetroffen. Het dikkopje bleef belangrijk als planktivore samen met de brakwatergrondel en zeenaald in 2011. In 2017 werden andere benthische soorten gevangen zoals de driedoornige stekelbaars en kooraarvissen en domineerde de planktivoren de aantallen en biomassa met soorten zoals het dikkopje, haring en sprot. In 2011 droeg de zeedonderpad, als bentho-piscivore, duidelijk bij aan de totale biomassa per vak. De zeedonderpad werd in 2013 niet gevangen maar wel éénmalig in 2017.

Figuur 9.25: Aandeel van biomassa (procent, links) en dichtheid (procent, rechts) voor elke voedselgilde in verschillende vakken in het Grevelingenmeer in voorjaar 2008, 2011, 2013 en 2017.
De visstand in het Grevelingenmeer wordt tegenwoordig gekenmerkt door redelijke aantallen maar een lage biomassa. De lage biomassa is te verklaren doordat het visbestand voornamelijk bestaat uit kleine vis. De soorten die nu met regelmaat in het Grevelingenmeer worden aangetroffen zijn het dikkopje, brakwatergrondel, zwarte grondel, haring, sprot, botervis, zeedonderpad en koornaarvissen. Dit beeld verschilt niet heel veel met de resultaten uit het onderzoek uit 1994 (Meijer 1995). Daarnaast is de soortenrijkdom nog steeds het hoogst in het westelijke deel van het Grevelingenmeer, zoals ook was aangegeven door Meijer en Waardenburg (1990), en neemt vervolgens af richting het oosten nabij de Grevelingendam. Echter Hop (2017) gaf aan dat het aantal soorten weer licht toeneemt nabij de Flakkeese spuisluis welke toen net was geopend. Rondom de Brouwerssluis en Flakkeese spuisluis kunnen soorten worden aangetroffen zoals de glasgrondel, zeedonderpad, schar, schol en steenbolk, maar ook haring en sprot welke deels worden ingelaten met het water uit de Noordzee of Oosterschelde (Hop 2017).
Samenvattend
De visfauna is niet zeer rijk, en is in sterke mate beïnvloed geweest door het sluisbeheer. Na de afsluiting zaten vissen opgesloten, was in- en uittrek niet meer mogelijk en verouderde de platvispopulatie. Na de opening van de Brouwerssluis (1978) en de jaarrond openstelling (1999) was seizoensgebonden trekgedrag weer mogelijk, m.n. uitwisseling van mariene juveniele soorten en zomergasten. De laatste jaren bestaat het visbestand voornamelijk uit kleine vis met een relatief lage totale biomassa tot gevolg. De soorten die nu met regelmaat in het Grevelingenmeer worden aangetroffen zijn het dikkopje, brakwatergrondel, zwarte grondel, haring, sprot, botervis, zeedonderpad en koornaarvissen. De soortenrijkdom is het hoogst in het westen en neemt naar het oosten toe af. De opening van de Flakkeese spuisluis (winter 2017) lijkt een positief effect te hebben op de visstand in het oostelijk deel van het meer echter zijn hierover maar beperkte gegevens beschikbaar.
Bron: WMR rapport C089/19 paragraaf 1.4.2.
Zie ook Deltaexpertise wiki: Ontwikkeling van de vispolulatie
NB: ** Glasaalmonitoring RAVON + waterschap, monitoring ** vis(sterfte), Tidal Test Centre ** Merkprojecten haaien en roggen ** Hengelvangstregistraties (MijnVismaat)
9.11 Vogels
9.11.1 Natura 2000
Onderstaande beschrijving is afkomstig van de internetpagina over Natura 2000, waar ook een overzicht te vinden zijn met alle broedvogel- en niet-broedvogelsoorten waarvoor het Natura 2000-gebied is aangewezen met bijbehorende doelen in de Grevelingen.
De Grevelingen is van uitzonderlijk belang voor visetende watervogels. Het heldere water speelt hierin waarschijnlijk een rol. Voor fuut en middelste zaagbek is dit het belangrijkste overwinteringsgebied in Nederland. Ook voor kuifduiker, dodaars, lepelaar en kleine zilverreiger is het gebied van grote betekenis, terwijl geoorde futen zich in de nazomer verzamelen tot een groeiende ruiconcentratie met internationale aantrekkingskracht en een voor Nederland verder ongekende omvang. Ook voor de brilduiker, benthos/viseter, is de Grevelingen het belangrijkste overwinteringsgebied. Terwijl de kleinere en kustgebonden viseters recent sterk toenamen, is het belang van de Grevelingen voor fuut, aalscholver, middelste zaagbek en brilduiker rond 1999 verminderd, mogelijk in samenhang met het gewijzigde sluisbeheer. Een minder gunstige situatie kan ook ontstaan door het optreden van stratificatie in de diepere delen, die invloed kan hebben op de visstand. Stratificatie is gerelateerd aan beperkingen in doorstroming en peilvariatie. Behalve voor viseters is het gebied verder van belang voor enkele ganzen, eenden en steltlopers, met name brandgans en strandplevier. Voor steltlopers die in de noordtak van de Oosterschelde foerageren is het gebied tevens van belang als hoogwatervluchtplaats. Kanoeten, die wat hogere eisen stellen aan hoogwatervluchtplaatsen (buitendijkse, verstoringsvrije schorren en ondiepten) overtijen bijv. bij Battenoord en Herkingen.Zeer belangrijk broedgebied voor kustbroedvogels van zandplaten en schelpenstrandjes (kluut, bontbekplevier, strandplevier, grote stern, visdief en dwergstern).
| Naam | Wetenschappelijke naam | Broedvogels | Winter- en trekvogels |
|---|---|---|---|
| Bontbekplevier | Charadrius hiaticula | x | x |
| Kluut | Recurvirostra avosetta | x | x |
| Strandplevier | Charadrius alexandrinus | x | x |
| Visdief | Sterna hirundo | x | NA |
| Dwergstern | Sternula albifrons | x | NA |
| Grote stern | Thalasseus sandvicensis | x | NA |
| Bruine kiekendief | Circus aeruginosus | x | NA |
| Goudplevier | Pluvialis apricaria | NA | x |
| Kievit | Vanellus vanellus | NA | x |
| Lepelaar | Platalea leucorodia | NA | x |
| Tureluur | Tringa totanus | NA | x |
| Slechtvalk | Falco peregrinus | NA | x |
| Aalscholver | Phalacrocorax carbo | NA | x |
| Bergeend | Tadorna tadorna | NA | x |
| Brandgans | Branta leucopsis | NA | x |
| Brilduiker | Bucephala clangula | NA | x |
| Fuut | Podiceps cristatus | NA | x |
| Grauwe gans | Anser anser | NA | x |
| Kleine zwaan | Cygnus columbianus | NA | x |
| Knobbelzwaan | Cygnus olor | NA | x |
| Kolgans | Anser albifrons | NA | x |
| Krakeend | Mareca strepera | NA | x |
| Kuifduiker | Podiceps auritus | NA | x |
| Meerkoet | Fulica atra | NA | x |
| Middelste zaagbek | Mergus serrator | NA | x |
| Pijlstaart | Anas acuta | NA | x |
| Rotgans | Branta bernicla | NA | x |
| Slobeend | Spatula clypeata | NA | x |
| Smient | Mareca penelope | NA | x |
| Wilde eend | Anas platyrhynchos | NA | x |
| Wintertaling | Anas crecca | NA | x |
| Rosse grutto | Limosa lapponica | NA | x |
| Geoorde fuut | Podiceps nigricollis | NA | x |
| Bonte strandloper | Calidris alpina | NA | x |
| Wulp | Numenius arquata | NA | x |
| Scholekster | Haematopus ostralegus | NA | x |
| Zilverplevier | Pluvialis squatarola | NA | x |
| Kleine zilverreiger | Egretta garzetta | NA | x |
| Dodaars | Tachybaptus ruficollis | NA | x |
| Steenloper | Arenaria interpres | NA | x |
9.11.2 Rode lijst
De rode lijst van voorkomende vogels in Nederland is terug te vinden bij Ministerie van Landbouw, Natuur en Voedselkwaliteit.
9.11.3 Voorkomen en trends van broedvogels en doortrekkers (SOVON)
Door SOVON Vogelonderzoek Nederland wordt de monitoring voor de Grevelingen (gebied 115) gepubliceerd. De figuren hieronder zijn direct verkregen van de SOVON website. Hieronder worden alleen de soorten weergegeven die langjarig zijn geregistreerd door de SOVON vogeltellingen.
Grevelingen SOVON gebiedspagina
Het kan zijn dat niet alle data die bij SOVON beschikbaar is in onderstaande overzichten meekomt. Voor de meest accurate stand van zaken betreft vogeltellingen verwijzen we naar de SOVON gebiedspagina’s.
Trends Broedvogels
Steltlopers






Meeuwen






Roofvogels


Trends Winter-en trekvogels
Steltlopers


























Roofvogels


Watervogels












































9.12 Zoogdieren
Methode
Van de zeezoogdieren zijn de gewone en de grijze zeehond onderzocht, beiden Natura 2000 soorten. De trendanalyses en beschrijving van de ontwikkeling van Natura 2000 vogelsoorten en zeezoogdieren zijn uitgevoerd door Delta Project Management in opdracht van Wageningen Marine Research. Onderstaande teksten zijn samengevat naar Arts, F.A.; Hoekstein, M.S.J.; Sluijter, M. (2019)Analyse en prognose trends vogels en zeehonden Grevelingenmeer. Delta Project Management. 2019-06. De data zijn afkomstig van Rijkswaterstaat Centrale Informatievoorziening. Vanaf 1987/1988 worden maandelijks alle watervogels, roofvogels en zeezoogdieren in het Grevelingenmeer geteld.
Resultaten
Na het openen van de sluis in de Brouwersdam in 1999 duurde het twee jaar voordat de eerste zeehonden ‘doorhadden’ dat er aan de andere zijde zich een visrijk water bevond. Vanaf 2008/2009 werden elk jaar weer meer zeehonden geteld. In 2017/2018 werden maximaal 70 exemplaren gewone zeehonden geteld. De aantallen die worden waargenomen per telling fluctueren sterk, vermoedelijk zwemmen de zeehonden regelmatig in en uit het Grevelingenmeer door de Brouwerssluis. Daarbij komt dat de zeehonden in het Grevelingenmeer onafhankelijk van het tij kunnen rusten en waarschijnlijk geen getijritme hebben zoals in de getijdewateren waar de zeehonden met laag water rusten op de droogvallende platen. Het is sterk afhankelijk van weersomstandigheden of de zeehonden op het droge liggen en dan goed zichtbaar zijn. De gebruikte methodiek is hierdoor minder geschikt om zeehonden te tellen en de getallen kunnen daardoor lager liggen dan de tellingen van Staatsbosbeheer. Daarnaast vindt regelmatig verstoring plaats waardoor de dieren het water in duiken. In het Grevelingenmeer rusten de zeehonden op de basaltdammen die dienen als oeververdediging. Groepen zeehonden worden met name gezien op de oeververdedigingen rond de eilanden Stampersplaten en Veermansplaat. De trend in het Grevelingenmeer volgt de trend in de Zoute Delta.
Net als de gewone zeehond heeft de grijze zeehond het Grevelingenmeer ontdekt. Maar die is daar nog een vrij zeldzame verschijning. In 2017/2018 werden maximaal 3 exemplaren waargenomen. De grijze zeehond komt massaal voor in de Voordelta maar niet in de overige deltawateren, dit in tegenstelling tot de gewone zeehond die ook talrijk is in de Oosterschelde en Westerschelde.
Samenvattend
De gewone (vele tientallen) en de veel schaarsere grijze zeehond nemen in het Grevelingenmeer toe als uitvloeisel van de sterke toenames in de getijdewateren van het Deltagebied. Dit proces zal waarschijnlijk nog enige tijd doorzetten, al is sterfte als gevolg van verdrinking in fuiken mogelijk een beperkende factor.
9.13 Exoten
Het Grevelingenmeer wordt gekenmerkt door een groot aantal exoten en sommige exoten spelen een belangrijke rol in het ecologisch functioneren. Bij de macrowieren zien we bijv. het Japanse bessenwier explosief ontwikkelen. Voor het eerst waargenomen in 1980, bereikt het areaal een maximum van bijna 60 ha in 1989. Daarna neemt het areaal weer af, de oorzaak hiervan is onbekend. Bij de bodemfauna zien we dat zowel muiltje als sinds de jaren ’00 de Japanse oester de biomassa van bodemdieren in sterke mate bepaald. Als rifvormende, epibenthische soort heeft de Japanse oester een invloed op de biodiversiteit doordat allerlei andere soorten op de oesters kunnen groeien en in de riffen allerlei mobiele soorten kunnen foerageren dan wel schuilen. Sinds 2005 wordt de Amerikaanse ribkwal in toenemende mate waargenomen in het Grevelingenmeer. De ecologische impact is onbekend, maar van andere gebieden is bekend dat de impact groot is, bijv. op het visbestand door het massaal opeten van vislarven.
9.14 Beggiatoa matten
Link naar Delta-expertise over Beggiatoa matten Delta Expertise: Grevelingen